#homozygous behaviour
Text
Calcium Phosphate Market: Global Industry Analysis and Forecast 2023 – 2030

The Global Calcium Phosphate Market size is expected to grow from USD 748 million in 2022 to USD 1113.58 million by 2030, at a CAGR of 5.1% during the forecast period (2023-2030).
Calcium phosphate is a common inorganic compound. It is a calcium salt of phosphoric acid and is widely used in variety of industrial areas, including the pharmaceuticals, and food & agriculture. The diverse applications of calcium phosphate in various industries are the major driving factor for the expansion of the market. Favorable properties such as higher nutritional value and extendable shelf life are to facilitate the growth of the calcium phosphate market. Fast urbanization, rising disposable incomes, dynamic change in consumer behaviour, and growing demand for nutritional food are hereby expected to stimulate the demand for calcium phosphate. Rise in the number of patients who are suffering from rickets, osteoporosis, homozygous thalassemia, osteomyelitis, fracture, will enhance the product acceptance in tooth & dental care sector. The demand for calcium phosphate in agriculture is expected to increase as it is an essential ingredient for manufacturing fertilizers.
Get Full PDF Sample Copy of Report: (Including Full TOC, List of Tables & Figures, Chart) @
https://introspectivemarketresearch.com/request/15824
The latest research on the Calcium Phosphate market provides a comprehensive overview of the market for the years 2023 to 2030. It gives a comprehensive picture of the global Calcium Phosphate industry, considering all significant industry trends, market dynamics, competitive landscape, and market analysis tools such as Porter's five forces analysis, Industry Value chain analysis, and PESTEL analysis of the Calcium Phosphate market. Moreover, the report includes significant chapters such as Patent Analysis, Regulatory Framework, Technology Roadmap, BCG Matrix, Heat Map Analysis, Price Trend Analysis, and Investment Analysis which help to understand the market direction and movement in the current and upcoming years. The report is designed to help readers find information and make decisions that will help them grow their businesses. The study is written with a specific goal in mind: to give business insights and consultancy to help customers make smart business decisions and achieve long-term success in their particular market areas.
Leading players involved in the Calcium Phosphate Market include:
Innophos(US), Prayon(Belgium), ICL PP (Israel), Chengxing Industrial (China), Hens(Indonesia), Budenheim (Germany), Tianjia Chem (China), Hindustan Phosphates (India), Thermphos (Netherlands), Kolod Food Ingredients (China), Gadot Biochemical (Israel), Haifa Group – (Israel), Advance Inorganics (India), Triveni Chemicals (India), Chemicals Chamber (South Africa)
If You Have Any Query Calcium Phosphate Market Report, Visit:
https://introspectivemarketresearch.com/inquiry/15824
Segmentation of Calcium Phosphate Market:
By Type
Monocalcium phosphate
Dicalcium phosphate
Tricalcium phosphate
By Application
Pharmaceutical
Foods & Beverages
Chemicals
By Regions: -
North America (US, Canada, Mexico)
Eastern Europe (Bulgaria, The Czech Republic, Hungary, Poland, Romania, Rest of Eastern Europe)
Western Europe (Germany, UK, France, Netherlands, Italy, Russia, Spain, Rest of Western Europe)
Asia Pacific (China, India, Japan, South Korea, Malaysia, Thailand, Vietnam, The Philippines, Australia, New Zealand, Rest of APAC)
Middle East & Africa (Turkey, Bahrain, Kuwait, Saudi Arabia, Qatar, UAE, Israel, South Africa)
South America (Brazil, Argentina, Rest of SA)
What to Expect in Our Report?
(1) A complete section of the Calcium Phosphate market report is dedicated for market dynamics, which include influence factors, market drivers, challenges, opportunities, and trends.
(2) Another broad section of the research study is reserved for regional analysis of the Calcium Phosphate market where important regions and countries are assessed for their growth potential, consumption, market share, and other vital factors indicating their market growth.
(3) Players can use the competitive analysis provided in the report to build new strategies or fine-tune their existing ones to rise above market challenges and increase their share of the Calcium Phosphate market.
(4) The report also discusses competitive situation and trends and sheds light on company expansions and merger and acquisition taking place in the Calcium Phosphate market. Moreover, it brings to light the market concentration rate and market shares of top three and five players.
(5) Readers are provided with findings and conclusion of the research study provided in the Calcium Phosphate Market report.
Our study encompasses major growth determinants and drivers, along with extensive segmentation areas. Through in-depth analysis of supply and sales channels, including upstream and downstream fundamentals, we present a complete market ecosystem.
If you require any specific information that is not covered currently within the scope of the report, we will provide the same as a part of the customization.
Acquire This Reports: -
https://introspectivemarketresearch.com/checkout/?user=1&_sid=15824
About us:
Introspective Market Research (introspectivemarketresearch.com) is a visionary research consulting firm dedicated to assisting our clients to grow and have a successful impact on the market. Our team at IMR is ready to assist our clients to flourish their business by offering strategies to gain success and monopoly in their respective fields. We are a global market research company, that specializes in using big data and advanced analytics to show the bigger picture of the market trends. We help our clients to think differently and build better tomorrow for all of us. We are a technology-driven research company, we analyse extremely large sets of data to discover deeper insights and provide conclusive consulting. We not only provide intelligence solutions, but we help our clients in how they can achieve their goals.
Contact us:
Introspective Market Research
3001 S King Drive,
Chicago, Illinois
60616 USA
Ph no: +1-773-382-1047
Email: [email protected]
#Calcium Phosphate#Calcium Phosphate Market#Calcium Phosphate Market Size#Calcium Phosphate Market Share#Calcium Phosphate Market Growth#Calcium Phosphate Market Trend#Calcium Phosphate Market segment#Calcium Phosphate Market Opportunity#Calcium Phosphate Market Analysis 2023
0 notes
Text
Fwd: Graduate position: UBern.Two.MouseInbreeding
Begin forwarded message:
> From: [email protected]
> Subject: Graduate position: UBern.Two.MouseInbreeding
> Date: 1 November 2023 at 04:41:30 GMT
> To: [email protected]
>
>
>
> Two PhD positions (4 years) on the effects of inbreeding on robustness,
> resilience, and replicability in mice
>
> At the Division of Animal Welfare (Prof. Hanno W�rbel, PD Dr. Bernhard
> Voelkl), University of Bern, Switzerland, we are seeking two PhD
> students to study how inbreeding affects developmental robustness
> and stress resilience in laboratory mice, and how this impacts the
> replicability of research findings and animal welfare. Both positions
> are part of a research project funded by the Swiss National Science
> Foundation (SNSF). The successful candidates will join a dynamic team
> of scientists and technicians involved in national and international
> research projects aimed to promote rigorous and responsible animal
> research. For further information on our team and research, visit our
> website (https://ift.tt/z1kPrsi).
>
> PhD student 1
> will analyse large public databases to investigate the effects
> of homozygosity on phenotypic plasticity and on environmental and
> mutational robustness in laboratory mice and conduct a systematic review
> and meta-analysis on the effect of inbreeding on stress resilience and
> the replicability of research with mice.
>
> PhD student 2
> will experimentally test whether homozygous (inbred) mice are
> developmentally less robust and less resilient to stressors by assessing
> morphological asymmetries and behavioural, physiological, and cellular
> measures of stress using a representative population of inbred, F1 hybrid,
> and outbred mice.
>
> Candidates need a university degree in biology or biomedical sciences
> and advanced training in statistics. Expertise in data analysis and
> quantitative genetics (for PhD 1), or morphometrics and experience
> with laboratory mice (PhD 2), will be a plus. We offer an attractive
> academic environment, opportunities for academic career development,
> and a competitive salary based on the Swiss National Science Foundation
> (SNSF) scheme.
>
> Please send your application letter together with a motivation statement,
> your CV, copies of relevant study certificates, and contact details of
> one or two reference persons (reference letters are not required at this
> stage) merged into one single pdf-file to: [email protected].
>
> The deadline for application is November 30, 2023. The position will
> be available from January 2024 or at your earliest convenience. Please
> indicate your preferred, as well as your earliest possible start date
> in the application letter.
>
> For informal enquiries, please contact Prof. Hanno
> W�rbel: [email protected] or PD Dr. Bernhard Voelkl:
> [email protected].
>
>
> Bernhard Voelkl
> Animal Welfare Division
> Vetsuisse Faculty, University of Bern
> Laenggassstrasse 120, CH-3012 Bern
> [email protected]
>
>
> "[email protected]"
0 notes
Text
Calcium phosphate Market Analysis, Growth, Shares, Size, Trends, Challenges with Forecast to 2028

The Calcium Phosphate Market was valued at USD 711.70 Million in 2021 and is expected to reach USD 988.16 Million by the year 2028, at a CAGR of 4.8%.
Calcium phosphate is a common inorganic compound. It is a calcium salt of phosphoric acid and is widely used in variety of industrial areas, including the pharmaceuticals, and food & agriculture. The diverse applications of calcium phosphate in various industries are the major driving factor for the expansion of the market. Favorable properties such as higher nutritional value and extendable shelf life are to facilitate the growth of the calcium phosphate market. Fast urbanization, rising disposable incomes, dynamic change in consumer behaviour, and growing demand for nutritional food are hereby expected to stimulate the demand for calcium phosphate. Rise in the number of patients who are suffering from rickets, osteoporosis, homozygous thalassemia, osteomyelitis, fracture, will enhance the product acceptance in tooth & dental care sector. The demand for calcium phosphate in agriculture is expected to increase as it is an essential ingredient for manufacturing fertilizers. Increasing awareness among farmers to improve crop quality and yield is raising the demand for the high in quality fertilizers and is estimated to boost the calcium phosphate market growth in the forecasted period.
The Research Report on Calcium phosphate Market provides a detailed overview of the demand and consumption of various products/services related to the growth dynamics of the market during the forecast period 2022-2028. In-depth market estimates of the various opportunities in the segment are expressed. Volume and revenue, Insights and analysis on the Calcium phosphate Market spans multiple pages. These are covered in various sections, including drivers and constraints, challenges and opportunities, regional segmentation and opportunity assessment, end-use/applicability forecast analysis, and competitive landscape assessment.
Read More: https://introspectivemarketresearch.com/reports/calcium-phosphate-market/
#Global Calcium phosphate#Calcium phosphate Size#Calcium phosphate Share#Calcium phosphate Growth#Calcium phosphate Trend#Calcium phosphate segment#Calcium phosphate Opportunity#Calcium phosphate Analysis 2022#US Global Calcium phosphate#Global Calcium phosphate Industry Size
0 notes
Text
My ‘unhealthy’ chickens
My blog has a lot of cute chicken photos on it, and sometimes I get comments such as “wow I want a chicken like that!” Which is lovely! I’m glad to spread the chicken love! However I want to take a second to address this. Just like with dogs, there are many breeds of chicken which are unhealthy and have health consequences because of this. I have a couple of birds like this. I just want to make people aware that if they seriously want a bird like this, to take into account what health issues may come with them.
Sooty - Frizzle feathers

Sooty is a fan favourite, and one of my favourites too. I do admit that I love the frizzle feather gene, it just looks so cool! However frizzle feathering causes some issues and I didn’t intentionally get a frizzle, Sooty hatched from a mystery egg. Her Dad was the only frizzle in the flock, with 13 other roosters, so she was a surprise to say the least!
Weather intolerance: Due to the feathers sticking out like that, birds can’t warm up in the cold since their feathers don’t provide a protective barrier and body heat escapes. This also means they have no natural shelter from rain and wind. Sooty doesn’t have to worry about any of this, it’s never cold where I live and she lives inside. She really struggles in the heat, however that’s likely due to her leg and foot feathering which I’ll discuss later, rather than the frizzle feathering.
Flight: Birds can’t fly well, since their primary wing feathers are curled or brittle. This puts them at risk of leg and spinal injuries if they try to fly from too large a height. Sooty did severe nervous damage to her spine when she was 8 weeks old, causing her to become paralysed in the legs for 3 months. Thankfully, she fully recovered after 5 months of physiotherapy.
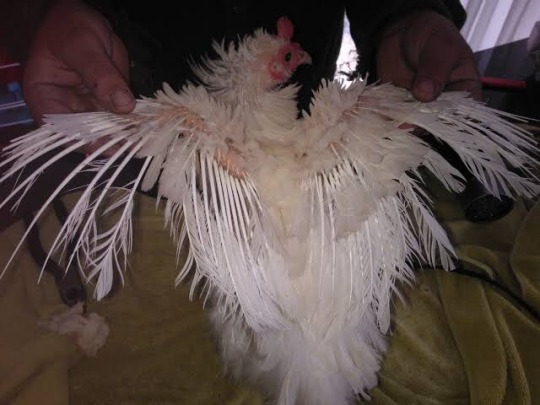
Sooty’s wings look like this. Not all frizzle feathered birds have flights this poor, however it is a potential consequence of the feather type.
Communication: Another issue frizzle feathers cause is communication within a flock. Sooty used to get pecked a lot by her top hen Kath, because Kath thinks Sooty is always challenging her to fight! Chickens use their neck feathers, called hackle feathers, to communicate a whole bunch of things. From fear, to aggression, to even asking another flock member to clean their feathers. Since frizzle feathers stick out like that, the bird struggles to move them into the positions used for communication so can’t talk to their flock very well. Sooty’s curled hackle feathers make it look like she’s always challenging another hen to a fight

You can see how those raised hackles kinda look like Sooty’s ‘mane’ of curled neck feathers! Thankfully, Sooty is second in command and her head hen, Kath, seems to have learnt that Sooty just looks like that! So she doesn’t get attacked very frequently anymore.
Unethical breeding: The gene which causes frizzle feathers in homozygous form (two copies of the gene) also causes serious issues. These birds are called Frazzles or ‘over frizzled’ and their feathers are very weak, often falling out and leaving the bird naked. It can be painful for them, and if they’re left outside, they certainly wouldn’t survive. Therefore frizzles should never be bred together. To breed frizzles you should use a frizzle feathered bird with a smooth feathered bird. However, this means you hatch only 50% frizzles, so some unethical breeders breed frizzle x frizzle to reduce the amount of smooth feathered birds they get.

A Frazzle chicken
Heart failure: These feathers have also been linked with enlarged hearts, increasing risk of heart failure. Since the feathers cause loss of body heat, it causes an increase in metabolism and other physiological functions to keep the body temperature at the appropriate level. This means the heart has to work harder, increasing its size and putting more strain on it. Sooty tires out easier, and when she used to be out ranging she’d frequently come inside to sleep on the couch while her flock was still outside having fun. I could see this putting them more at risk of predation, since if they’re already tired they don’t have the same stamina of another bird to flee a predator.
Solo - Heavy foot/leg feathering

Not the most flattering photo of her but the best one I have to show her foot feathering! As you can see she was quite cranky with me! She’s a Silkie X Pekin, which are both breeds known to have heavily feathered legs. There are many breeds with healthy foot feathering, such as Langshans

But some like the show-type Pekin bantam, have a number of issues associated with their foot feathering. Here’s a Pekin in comparison to the Langshan above
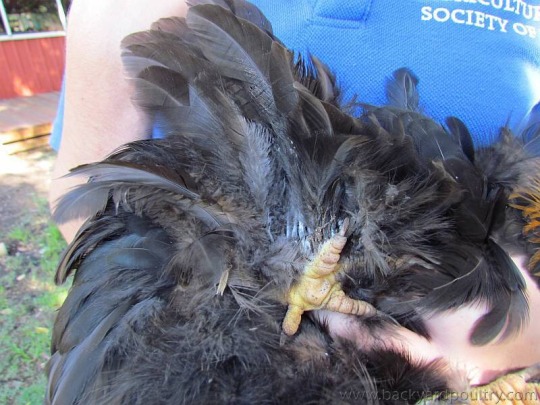
As you can see the feathering is much much heavier!
Mobility: Very heavy leg and foot feathering significantly reduces mobility. The large feathers make it harder to move toes, making perching more difficult, and are a tripping hazard. Solo is always tripping over, stumbling, and ‘shuffling’ when she walks since her feet impair her movement a significant amount. I’m probably going to cut her foot feathers off so she can move about easier. They’d never hindered her movement until now, this molt they grew in humongous for some reason. Obviously having a built in tripping hazard isn’t a good idea, since it predisposes the bird to a higher risk of leg injuries.
Thermotegulation: As mentioned above, legs and feet are very important in helping a bird regulate their temperature. Lightly feathered legs like the Langshan has don’t have this issue, since the bird has majority of its foot free to cool down with. Heavily feathered legs like the pekin provide little surface area to cool down with, so the birds can really struggle in hot weather. Solo is one of our least heat tolerant birds, and she thankfully has wattles and a comb unlike poor Silkies!
Cleanliness: Heavily feathered feet get disgusting! They’re more prone to getting dirty and are harder for the birds to keep clean. Solo always has poop, sticks, food, mud, and all sorts of other gunk crusted into her foot feathering. I have to clean them quite frequently so that she doesn’t get bacterial build up.
Other health ailments: In my experience, heavily feathered feet tend to be a beacon for related leg and foot health issues. We don’t have to worry about this where I live, but foot feathers can get wet in snow and heighten frostbite risk for toes. Although I don’t have to worry about the cold, sadly these foot feathers also have heat related issues! I live in a sub-tropical environment, so humidity levels get pretty high here. Bacteria loves humidity. Solo has had a bad case of Bumblefoot which was really hard to treat due to this humidity. Sweep, another bird with heavy foot feathering, has had 2 cases of bumblefoot now. I’ve never had a clean-legged bird get bumblefoot, so it’s definitely linked to trapping bacteria and humidity. I haven’t had to deal with this parasite myself, but apparently feather-legged breeds are more prone to Scaly-leg mite too.
Cujo - Heavy layer breeds

Cujo is a Hamburg, sometimes referrred to as ‘Everlayers’ since they have a reputation for reliably laying an egg every day. They rarely go broody, and if they do are very easy to dissuade. I am very against production breeds if they’re not within an agricultural industry, where they have a purpose, since it tragically shortens their lives so much. The Hamburgs lay on average 200 eggs annually, which isn’t too bad and makes them a healthier layer breed, but it’s certainly heavier laying than most of the other breeds I have. Cujo is actually very healthy, I took great care in picking a breeder to get her from and most of his birds are lighter layers than they ‘should’ be. Cujo was laying 3-4 eggs a week before her current molt, much better than the 5-7 her breed has a reputation for.
Heart failure: One of the most frequent ends to laying birds is heart failure. Their bodies are under so much stress to make an egg every day that their bodies eventually just give up, usually from heart failure.
They don’t go broody: A lot of people don’t like broody hens, since they stop laying and sit on their nest all day, however I really like them. A broody hen gets a much needed break from laying eggs! Some breeds continue to lay eggs over winter, and some birds don’t stop laying when they molt if it’s a light one. So broodies give the bird a choice to stop laying and sit on eggs when she wants, if she didn’t get a break over winter or molt. Breeding this behaviour out of production breeds contributes to their issues, since they can’t take that break.
Shortened life span: Due to the strain mass egg production puts on their bodies, average lifespan is 3-4 years compared to the 6-10+ of healthy heritage breeds. I had a utility leghorn as a pet many years ago, her heart tragically gave out on her one day while I wasn’t home. She was dirtbathing in her favourite spot when it happened, so I hope to think it was a peaceful end. She was only 2 years old.
Reproductive complications: Heavy layer breeds are more prone to experiencing issues with their reproductive tract. This includes cancers, tumours, prolapses, egg binding, and egg yolk peritonitis (infection). They’re also more prone to nutrient deficiencies, especially with calcium, since it takes so much out of them to lay eggs. This is easily preventable with a balanced diet, however if calcium deficiency does occur the hen can suffer from brittle bones.
Sweep - Aggressive breeds

Now Sweep isn’t nearly as bad as this title frames her to be, but it’s still worth a mention. We can only guess what her parentage is since she came from mystery eggs, but we think Sweep is an Old English Game cross Pekin. In Australia, Old English Game are a hyper aggressive breed. They were bred for the cruel sport of cockfighting, where two roosters are forced to fight to the death. Thankfully this sport is now criminalised, but nobody bred the aggressiveness out of this breed.
Injury risk: hyper aggressive breeds pose a greater risk of injury mainly to other flock mates, but also to themselves. These birds often antagonise others despite there not really being a reason to, resulting in more fighting, disharmony, and injury within a flock. If another bird is stronger than them and gets sick of their shit, they themselves could be seriously hurt since they often don’t know when to back down from a fight like non-aggressive birds do. Sweep has to be housed separate from my main flock with her mother, Solo for company. She has tried many times to outrank birds in my main flock but her fighting is very brutal compared to the normal pecking order fights. She aims for the eyes, and came close to blinding a bird once before, I can’t risk that sort of injury.
Mortality risk: continuing on from that first point, some individuals will take their aggressiveness too far and kill fellow flock mates. What might start out as a simple pecking order fight can turn very bloody and very brutal with these breeds fast. Roosters can kill hens and hens can kill hens. This obviously should never happen in well bred, good tempered birds. I do not say this jokingly when I say that Sweep and Sooty would kill each other if I let them. They’re both Pekin X Old English, and although Sooty is good with other birds, she’s terrible with Sweep. I’m hoping Sweep will mellow with age (she’s currently 2) and I can integrate her and Solo as part of Blossom’s flock.
Social interaction: I think this is something a lot of people don’t seem to consider, but having hyper aggressive birds which have to be housed seperate will obviously hugely impact upon their social needs. It doesn’t matter how aggressive the bird is, a chicken is, and always will be, a social animal. They need companionship, and while this can be provided by us, it’s easiest to provide it with other chickens. Keeping a social animal by itself, never letting it interact with others, and not providing that companionship yourself is incredibly inhumane in my opinion. It doesn’t matter if that bird is incapable of interacting without trying to kill the others, the fact is that this animal is still hardwired to live in a social group. By breeding such aggressive animals, it’s very cruel since it deprives them of such a basic need.
Now this post isn’t to say people can’t get a breed if they like it but it has health consequences, because something like those heavy foot feathers don’t cause the bird any harm or pain in itself, it’s just a consequence of poor management. So if you’re willing to do the work to ensure those features don’t hinder the animals quality of life, then excellent! Go ahead and get those basketball-shaped Pekin lads! This post was merely a reminder to think critically and research any animal/breed before you get them, and to make sure you’re prepared for any future consequences or adjustments for that animal/breed. Sooty and Solo need fans set up on their pens during the Summer, Sweep needs a seperate coop, and all three need adjusted perches and weekly foot health checks because of their heavily feathered feet. Once again, the importance of you screening for ethical, responsible breeders is crucial when deciding to bring a new family member home.

Thanks for reading!
4K notes
·
View notes
Text
My Episode 7 Predictions!
Mother of God! How have these weeks passed by so fast?! I've decided to share what I think will be answered tomorrow night and some of the fates of our beloved characters! The series has gotten off to an even bigger success than it's predecessor and EVERYONE'S talking about it! Without further ado, let's see what I'm predicting!
Who's H/fourth man (or woman) and will we find out?
The question that's on the nation's minds. That has been boggling us since 2017... My personal prediction still stands as CC Philip Osborne, however... I think he's purely sitting on his arse and pulling the strings, whilst others are doing his dirty work. Those people are Patricia Carmichael: who in all honesty I'm manifesting isn't actually bent but is so career driven and a bit of a lick arse that she's just taking orders and not questioning anything, plus she doesn't think much of Ted either... I have joked that she's been shagging Osborne on the side which would be quite funny. I also believe Buckells has been Osborne's true scape goat for years. Someone as laid-back and unpassionate as Buckells is easily manipulated - I mean how the hell has he become a super? As Steve rightfully says, he couldn't order a piss up in a brewery. Seeing the connections go well back into the early noughties whilst Buckells was part of an oasis tribute band, it's an easy conclusion to make. Lastly I will mention Thurwell, now I think Thurwell may have been more involved on his own doing anyway but I think he'd still been pretty busy doing things on behalf of Osborne up until his alleged death in sunny Spain (Belfast but shh). I know a lot of people still think Buckells, some reckon pas-agg Pat is the top woman... and a few still think our beloved Kate is 'H'.
Now the other part of my point is actually finding out... I think us as viewers will certainly know the identity of this mysterious individual. But, I think we'll find out in the very last few seconds and poor AC12 won't be any the wiser which will open up the foundations for a possible series 7. I think a lot of things will be answered for us viewers but not everything for the characters. I know that if they had a series 7 there would be an other primary focus but I just think it would be clever. Just like how we found out as viewers at the end of series 1 that Dot was a wrong'un.
Is Thurwell really dead?
I'm still very amazed by the whole induction of Jimmy Nesbitt as a series of photos. I know we were supposed to have a secretive guest actor appearance and I believe it was Robert Carlyle's name floating about which would have been insane! But that moment we saw Jimmy, I yelled at the screen. So, this answers the question of seeing a character that had only been mentioned previously - which has been very much hiding in plain sight. Even with a rewatch, I had forgotten about Thurwell and therefore hadn't even considered him. But, I did mention the list from series 3 which did loosely tie in. Anyway, when we learned that Thurwell and his Mrs were now dead and clearly had been for a while - it was cleverly shot so we didn't see who it was, only the Spanish Police's word. You know, I think if we were to get a series 7, Thurwell could appear and he's in fact not dead. It's very rare you get a very well known actor in for something so small without it leading to bigger things. Although, they got Andi Osho, a very much loved comedian in purely for archival photos and videos... I do feel that Thurwell could have easily been on the other end of the fake MSN, and doing Osborne's dirty work - especially with the Spanish connection. In a BTS photo we have seen a clue to a piece of paperwork regarding a time share (Spain mentioned) in front of Kate which will likely have a connection to Thurwell and Spain. I think he's possibly still alive but we'll see...
Joanne Davidson
Kelly Macdonald has been welcomed with a very warm reception. Wow, her connection to Tommy Hunter (albeit disturbing, homozygous DNA... nasty business) was not something I ever expected although I guessed very much so when we learned of the DNA match with a nominal. Would I define her as bent? I mean in layman's terms, yes she is. However each guest star has had a very complicated relationship with the definition - all have done dodgy things and usually for good reason... But I do sympathise with her because of her background, born into trouble, it was very hard to escape - even if her mum took her to Scotland for a life away from it all. From the very start of her career she was doing the OCG's bidding and it transpires she was in the police on Tommy's orders - so very much another caddy. I do feel Jo had always wanted to be a good person and do right and in some ways she did - however she's in too deep. Next I'll mention her relationship with Kate and it's something I'd never have predicted. A lot of suggestive and sapphic behaviour from the off. Now, I do think Jo cares deeply for Kate and has developed strong feelings - and I think Kate deep down has also but I don't think anything will happen sadly. And that's more for the fact that they had run out of track before they could even get going as so much has happened. Kate's gonna save Jo when her transport gets ambushed, that's a given... but I don't think we'll see anything happen that we want to happen. I hope I'm very wrong and we get something but with the hour we've got I don't think it'll be the case. And as for her fate? I think she'll survive. Guest leads usually die, eventually... with the exception of Roz who's serving time with one arm in Brentiss. Giving how unfortunate her life has been from her conception, I think it'll do her justice to escape with her life. Witness protection? Hmm perhaps... Although I think she's done enough dodgy stuff to warrant a prison stretch. Will she be instrumental in revealing who the fourth dot is? Well yeah actually, I think she grew up thinking Thurwell was her dad... and of course having connections to both Osborne and Buckells. Jo joined the force just before the turn of the millennium so would have been a copper when the Lawrence Christopher case occurred in 2003. Even though not directly working with them, I'm sure she knew what was going on and would have come across Osborne and Buckells at that time. I have a strong inkling that she knows exactly who she's been talking to and has been under their wing since the start. If she dies I will be gutted.
Kate
Where are we going with Kate? Well she's just killed Ryan (lawfully) and has gotten away with it. But Carmichael isn't thick... and nor is Steve and Ted. They all know but I think that'll be that for now. For ages I thought she was gonna D word but I think she's gonna be alright now. I hope that somehow she gets back into anti-corruption permanently because that's where she's best suited. I'm shocked that she hasn't had a glass box spectacular - my predictions for her to be accused of being bent and all that were way off the mark... and not a single mention of the two dying declarations... With only an hour tomorrow and so much to deal with, I guess that's not going to be mentioned. And Kate ain't gonna be sanctioned either. But, there's always opportunity with a series 7... She'll definitely be back in AC12, interviewing... I can't see all the BTS photos being Jed Herrings. But we'll see tomorrow!
Steve
Poor Steve eh? He's not had an easy time of it. Addicted to painkillers as I rightfully predicted, unable to trust his gaffer, torn on a transfer and a up and down friendship with his 'mate'. Oh and the car, actually let's not talk about the Mazda... Anyhow I believe that Steve will talk about his back and recent struggles with meds after a lot of emails from occupational health. Will he get pulled off the job at the last second because he didn't book an appointment... yeah I think that might happen and Kate takes his place or something. But there is a counsellor or something credited so I'm pretty certain on that. Will he lose his long overdue promotion? It's possible! I don't think the test was going to come back as a fail but more of a cause for concern - just that the levels of codeine etc in his system was higher than it should and it would be best advised to talk to someone. We haven't seen much pill taking going on since the drugs test so has Steve been going cold turkey? Or have they just veered away from that part of the story? In all fairness he's appeared fine with his back since the Windermere convoy - he ran across an industrial estate... I mean. I hope Steve gets a glimmer of hope and happiness because he rightfully deserves it. Ever since the start he's had his fair sharing of aggro and he deserves a break. I do think however he may be partly responsible for nailing the final nails into Ted's coffin - especially after he discovered the 50K up Merseyside and the truth from Lee Banks. I think he could be potentially happy with Steph, it would be nice for something to happen with them but we'll see. Steve needs a big hug.
Ted
This is going to pain me saying it. But, I don't think Ted is going to make it to the end. His retirement is inevitable. I can see the argument of the retirement being rescinded if they unmask the fourth dot etc and solve stuff but realistically, he's past retirement age. He's so focused on getting to the end of this marathon that I don't think he will see it through and know who it is - because he'll... yeah I don't need to say it. How? I've been saying his heart is gonna pack in. He's going to be in deep trouble over that 50k because Steve and Kate know the truth. But why would the likes of Carmichael find out... Well we've seen a clue of 'definate' on some paperwork - I reckon Ted by coincidence also makes this spelling mistake often as it is one of the most commonly misspelt words and therefore will be in the 'H' running again - he wont be 'H' as I reckon Osborne makes the same mistake... He alleges he misspelt it cos he studied the texts carefully but I think he spelt it how he usually would. They'll all go down the wrong path and accuse him which will lead to his ultimate downfall. Another little teaser is Steve appearing to listen to Ted's 2019 glass box spectacular however notably Carmichael says AC12 interview and not AC3... so mixing two different interviews to tease us? Maybe Ted might feel he has no choice but to sacrifice his career for the truth. However it all gets too much. Lies cost lives... I can't see Ted going on from beyond here and Ted was always supposed to be a minor character until he was very well received by the audience and he was made a main character. Realistically, his story is told and he's at his endgame. I hope he survives, by god I do... He's one of the most loved fictional characters at present. But I have a really bad feeling and I'm worried! It seems a total Jed thing to do for Ted to meet his end without ever finding the truth... seeing that's what his heart has been set on for about four years... I'm sorry to even be going there but it's what I think will happen. And I want to be wrong.
Miscellaneous Predictions
I want to round this all off with other little points. Carmichael won't be bent, maybe still involved with anti-corruption, you love to hate her, there's so much more they could do with her character. I don't think Steph's dodgy at all, she's just a widow whose been helped out by Ted in an unlawful way. I think her and Steve would be well suited. I hope Chloe will get to the end and continue on in anti-corruption, she's got a lot of potential. If Osborne isn't sussed, I reckon he'll still be CC. DCC Wise isn't bent either, just getting on with her job. But if Osborne does get caught and I'm wrong, there will be a lot of chaos for both him and Wise. Farida gets out of jail and starts afresh. Buckells I reckon will get out but won't be apart of the police force anymore. Lomax isn't bent, just a regular cop trying to get the work done. The rest of Jackie's remains will be found under the workshop floor, maybe with someone else? God knows who. Or maybe with some evidence that helps lead AC12 on the right track. Steve will still be an officer and Kate will be back where she belongs. There's probably more but my head hurts.
Will there be a series 7?
Yes.
Thank you all for reading my jumbled thoughts all packed into a text post. I'm nervous but excited for tomorrow night's finale. Let's see if I get anything right... probably not...
#line of duty#6x07#my finale predictions#oh god people will not like some of these#kate fleming#steve arnott#ted hastings#joanne davidson#philip osborne#ian buckells#patricia carmichael#ted better not d word#thanks for reading#my predictions#martin compston#vicky mcclure#adrian dunbar#bbc#jed mercurio#h#i've probably forgotten stuff#but oh well a lot to cover#lies cost lives#series 7 please#if i've missed anything#always ask my thoughts on the thing i've missed#line of duty spoilers
16 notes
·
View notes
Note
I would unironically love to hear more about your research proposal?? It sounds fascinating and I'm hoping to go into neuroscience myself so this is right up my alley. How did you get into this field? What's your research methodology like? What are its real-world applications? Sorry if this is super upfront, I'm just really interested in your research!
Oh my God I was going through my asks and suddenly realised I completely forgot to answer this. Anon I’m so sorry I hope you’ll still read this. Okay so I can’t say too much because this research will actually be carried out at some point but I’ll try to go into detail as much as possible. This will probably be long so I’m going to put it under a cut.
The research project was part of a course called Research Methods I took last period. So what we want to do is see if dopaminergic axons lead serotonergic axons to the forebrain, since it’s a very important neurotransmitter system. The method can be divided into three aims:
1. Use TRAP-sequencing to label mitochondria in dopaminergic neurons, pull them out of the brain and do mass spectometry to identify proteins that are produced in the dopaminergic neurons.
2. Match the found proteins to mTOR pathway, a pathway that’s important for dopaminergic axonal growth.
3. Create genetic knockout mice where we remove the genes that code for the identified and matched proteins. There’ll be wildtype mice, heterozygous knockouts and homozygous knockouts.
TW: Use of mice as test animals.
We will then cut out the prefrontal cortex of the brains of mice in the early postnatal stages. The mice will be tested for medial prefrontal cortex innervation, using immusostaining for the dopaminergic and serotonergic axons. The brains will also be sent out for HPLC-analysis, so we can see the effect the gene removal has had on dopamine/serotonin neurotransmitter balance.
Remaining mice will be allowed to grow into adults and will be used for behavioural testing, where tests related to autism spectrum disorder will be used to asses any behavioural deficits. The brains of these mice will then be analysed in the previous ways to check for any long lasting connectivity and neurotransmitter issues.
The results will be relevant to quite a lot of neurodevelopmental disorders (autism, major depressive disorder, schizophrenia, etc) since they involve dopamine/serotonin neurotransmitter imbalances.
I’m so sorry for taking so long to reply, anon. I really hope you got to read this!!
2 notes
·
View notes
Text
Thelytoky in the honey bee
Review article
Thelytoky in the honey bee
Frances GOUDIE, Benjamin P. OLDROYD
Behavior and Genetics of Social Insects Laboratory A12, University of Sydney, Sydney, NSW 2006, Australia Received 24 July 2013 – Revised 11 November 2013 – Accepted 2 December 2013
Abstract – Thelytoky, the asexual production of females, is rare in honey bees. However, it is ubiquitous in workers of the Cape honey bee Apis mellifera capensis. Thelytoky allows some workers to be reincarnated into the queen phenotype, and thereby selects for reproductive competition among workers. Thelytoky also acts as an exaptation for the emergence of reproductive parasites, the most extreme example of which is an entirely clonal ‘cancerous’ lineage of workers (the Clone) that lethally parasitises colonies of another subspecies Apis mellifera scutellata. The Clone is an enigma because thelytoky results in the accumulation of homozygosity at any loci that are free to recombine, yet the Clone retains considerable heterozygosity. The Clone pays a cost for its thelytoky: the selective removal of homozygous offspring at each generation. We propose that workers, queens and Clones have differing abilities to endure the costs and benefits of sex and asexuality, accounting for the heterogeneous distribution of reproductive strategies across the A. mellifera capensis population. We further suggest that multiple factors must fall into place for thelytoky to emerge as an effective reproductive strategy in a honey bee population, and that geographic isolation resulting in genetic drift and founder effects may have enabled thelytoky to emerge in A. mellifera capensis. Finally, we consider the honey bee in the broader context of haplodiploid Hymenoptera, and argue that constraints on the evolution of sex in non-haplodiploid taxa may make sexual reproduction an evolutionary ‘one-way street’.
Apis mellifera / Apis mellifera capensis / asexual / thelytoky / reproductive parasitism
1. INTRODUCTION
In the typical image of a honey bee (Apis mellifera) colony, there is a queen reigning over her worker force of daughters with an iron… wing? The queen, and only the queen, lays eggs. If she chooses to fertilise an egg with stored sperm, it develops as a diploid daughter, a future queen or worker. Alternatively, if the queen lays an unfertilised egg, it develops into a haploid male, a drone that will eventually fly out and attempt to mate with virgin queens of other colonies.
This image might come close to approximating a particularly well-behaved colony of the European
Corresponding author: F. Goudie, [email protected] Manuscript editor: Stan Schneider
honey bee. However, in reality, like the human suburbs of the 1950s, even the best-behaved honey bee colonies can have nefarious goings on beneath the surface. Here, we review one of the most fascinating ways in which reality differs from outward appearance: the asexual production of diploid females via thelytokous parthenogenesis. We discuss the physiological, evolutionary and social consequences of thelytoky in the subspecies in which is best characterised, the Cape honey bee Apis mellifera capensis (hereafter, Capensis). We further discuss the possibility of thelytoky in other honey bee species and subspecies, and explore how thelytoky may have evolved in honey bees.
1.1. Thelytoky
Honey bees are haplodiploid. Diploid females are normally produced sexually, from fertilised
eggs, while haploid males develop from unfertil- ized eggs via arrhenotokous parthenogenesis. Both queens and workers are capable of laying unfertilised, male-destined eggs, although in most circumstances workers rarely utilise this ability (Visscher 1989; Winston 1991; but see Barron et al. 2001). Thelytoky is an alternative develop- mental pathway for the unfertilised egg, which results in the production of a diploid female offspring.
Thelytoky, the asexual production of females, is rare among animal taxa, where sexual repro- duction predominates (White 1984; Suomalainen et al. 1987). Examples of notable thelytokous animals include the anciently asexual bdelloid rotifer, which has gone without sex for millions of years (Mark Welch et al. 2004; Gladyshev and Meselson 2008) and the Amazon molly (Poecilia formosa), in which females must mate with males of another species before they can reproduce thelytokously. This odd behaviour causes local extinctions as molly females ‘steal all the men’ (Tiedemann et al. 2005; Heubel et al. 2009).
Thelytoky has evolved at least 255 times in populations of normally arrhenotokous haplodiploids (Normark 2003; Engelstadter 2008). Many transitions from arrhenotoky to thelytoky are driven by maternally transmitted endobacteria, such as Wolbachia, Rickettsia and Cardinium (Zchori-Fein et al. 2001; Huigens and Stouthamer 2003; Hagimori et al. 2006; Engelstadter 2008). One mechanism by which these bacteria drive their own propagation is by inducing female-producing parthenogensis to reduce or eliminate the production of males (a genetic dead end for the bacteria) by their host. However, there are a rapidly increasing number of examples of genetically determined thelytoky being identified in haplodiploids. In particular, the ‘molecular natural history’ movement (Keller 2007) is revealing a fascinating array of novel reproductive systems that are based on genetically determined thelytoky. While the ants have thus far yielded the greatest diversity of unusual reproduction systems based on thelytoky (e.g. Pearcy et al. 2004; Ravary and Jaisson 2004; Fournier et al. 2005; Gruber et al. 2010), the bees, and particularly the honey bees,
are beginning to show that they can be equally weird (Sumner and Keller 2008).
2. APIS MELLIFERA CAPENSIS
2.1. Thelytoky in Capensis
Thelytoky in bees was first identified in Capensis (Onions 1912). In this South African subspecies of honey bee, thelytoky is almost ubiquitous in workers (Verma and Ruttner 1983). When Capensis workers lay unfertilised eggs, the eggs usually develop into diploid female offspring via automictic thelytoky with central fusion (Verma and Ruttner 1983; Figure 1). In automictic thelytoky, the reduc- tional division of Meiosis II occurs as normal, resulting in four haploid nuclei. Diploidy is then restored by one of several mechanisms, each with a different genetic outcome (Pearcy et al. 2006). In Capensis, diploidy is restored by central fusion; the fusion of two non- homologous pronuclei as if one of the nuclei acted as a sperm. In the absence of meiotic recombination between a locus and the centro- mere, central fusion results in clonal reproduc- tion so that the genotype of the daughter is identical to the genotype of the mother. However, when recombination occurs, hetero- zygosity can be lost, so that the daughter will be homozygous for one of her mother's alleles (Suomalainen et al. 1987).
If a Capensis worker produces a daughter queen via thelytoky, she is genetically reincarnated in the form of a queen with no frog kissing required. This is no doubt why Capensis workers target their egg laying around existing queen cells, in places where queen cells are likely to be built and during periods of queen rearing (Figure 2). Around 40–60 % of queens produced during swarming events are the daughters of workers (Jordan et al. 2008; Allsopp et al. 2010). Thelytokously-produced Capensis queens go on to mate and reproduce sexually (Beekman et al. 2011). Capensis workers also utilise thelytoky to raise a replace- ment queen whenever they are queenless and broodless (Holmes et al. 2010).
Without recombination With recombination
iii
i
Mother
Meiosis I
Meiosis II
Central fusion
ndomisation of leles during combination
ii
Maintenance of
Daughter
chance of loss of
heterozygosity (AB) heterozygosity
Figure 1 Automixis with central fusion. Meiosis occurs as normal resulting in four haploid pronuclei (i). Pronuclei occupying the central position fuse for form the diploid zygote (ii). As this fusion is central, the pronuclei involved are descended from the two different homologous chromosomes (iii). In the absence of recombination, heterozygosity in the mother will be maintained in the daughter. When recombination occurs, alleles are randomised among the four pronuclei and as a result there is a 1/3 chance that heterozygosity will be lost in the offspring. This is allocation of alleles to the central pronuclei is an example of sampling without replacement. If one of the central pronuclei carries an A allele, there is a 1/3 chance that the other central pronucleus will carry the second A allele, and a 2/3 chance that it will carry one of the two B alleles. If you do not believe us (many readers will not) try writing ‘A ‘on two bits of paper and ‘B’ on two other bits. Draw one piece of paper at random: this is the first central pronucleus. Let us pretend it is an A. Now, what is the probability that the second pronucleus you draw will also be an A?
Thelytoky dramatically increases the repro- ductive potential of the honey bee worker, resulting in competition between workers and worker patrilines (lineages of full-sister workers, sharing a father) (Moritz et al. 1996; Figure 3). This tendency has selected for traits related to reproduction and reproductive com- petition in Capensis workers (Greeff and Villet 1993). Capensis workers often have a well- developed spermatheca (a sperm storage organ found in queens), which is absent in workers of other honey bee subspecies (Hepburn and Crewe 1991; Phiancharoen et al. 2010). Furthermore, the Capensis worker has an
average of 10–20 ovarioles per ovary (Ruttner 1977; Hepburn and Crewe 1990; Allsopp et al. 2003; Goudie et al. 2012a). In contrast, workers in arrhenotokous honey bee populations typi- cally have far fewer ovarioles (Amdam et al. 2004; Oldroyd and Beekman 2008). Ovariole number in worker patrilines is heritable and highly variable (Goudie et al. 2012a). This suggests that certain patrilines dominate repro- duction in Capensis.
The Capensis worker does not always limit herself to competing with her sisters over the production of new queens. Capensis workers are able to act as non-natal reproductive
Figure 2 Worker laid eggs in a queenless Capensis colony. i Egg laying workers focus around holes in the comb where queen cells are most likely to be built. ii Worker laid eggs on the outside of an existing mature queen cell. Photos by B Oldroyd.
parasites, entering foreign colonies and laying eggs that may be raised as queens. Non-natal workers are responsible for the production of between 0.5 and 46 % of new queens (Jordan et al. 2008; Allsopp et al. 2010; Holmes et al. 2010; Moritz et al. 2011). Variation in the degree of parasitism experienced by different colonies suggests that parasitism may be assisted by beekeeping methods (Dietemann et al. 2006a; Härtel et al. 2006). However, Holmes et al. (2010) observed rates of parasit- ism that were independent of apiary layout and distance between colonies. Furthermore, Neumann et al. (2001) found that Capensis workers disperse significantly more than other subspecies of A. mellifera and are more likely to parasitize queenless colonies. Whether or not
movement of workers between colonies is an active process, as it seems to be in bumble bees (Blacher et al. 2013) and stingless bee queens (Wenseleers et al. 2011), remains open to question. However, it appears that once a non-natal worker enters a nest, she targets queen cells for oviposition. In colonies with high rates of parasitic queen production (38 %), only 6.9 % of the workers were non- natal (Jordan et al. 2008). Thus, the reproduc- tive output of non-natal workers is disproportionally high, as is seen in colonies of the Asian species Apis florea (Nanork et al. 2005; Chapman et al. 2009) and Apis cerana (Nanork et al. 2007). This suggests the exis- tence of specialised parasitic genotypes within the Capensis population.
0.75
0.325
Arrhenotoky Thelytoky
Figure 3 The relatedness (r) of a focal worker (red circle) to other individuals in a honey bee colony. Females are represented by circles and males by squares. The queen wears the crown, however, all females have the potential to be raised as a queen (although subfamilies differ in their likelihood of doing so). In a colony in which workers reproduce arrhenotokously, the focal worker is more closely related to the son of her mother (r = 0.25) than the son of her half sister (r=0.125) and so selection favours policing behaviour to suppress the reproductive efforts of other workers. In a colony in which workers reproduce thelytokously, the focal worker can produce daughters that are related to her by unity (r=1). She can use this ability to produce daughters that might become queens, resulting in her effective genetic reincarnation as a queen. She is equally related to her sisters as she is to the thelytokous daughters of her sisters. In the same way, the queen is equally related to her daughters as she is to the thelytokous daughters of her daughters, and so it makes no difference to the colony as a whole if the new queen is produced sexually by the original queen or thelytokously by a worker. Therefore, selection for policing behavior is relaxed relative to arrhenotokous populations.
2.2. The Capensis Clone
By liberating the worker from reliance on a sexual queen, thelytoky has enabled the emer- gence of entirely asexual lineages of social parasites. On at least three occasions, two historic and one current, parasitic lineages have emerged as specialised reproductive parasites of the strictly arrhenotokous subspecies Apis mellifera scutellata (hereafter Scutellata; Martin et al. 2002).
While Capensis is confined to the southern- most tip of South Africa, Scutellata occupies the rest of the southern and most of central Africa (Hepburn and Radloff 1998; Figure 4). In 1990, a beekeeper moved approximately 200 commer- cial Capensis colonies across the stable hybrid zone that separates the two subspecies (Beekman et al. 2008; Allsopp and Crewe 1993) and into Scutellata range (Allsopp and Crewe 1993; Neumann and Moritz 2002). From here, Capensis workers drifted into (or perhaps
invaded) the local Scutellata colonies, com- menced laying and produced thelytokous daughters. One of these daughters founded a thelytokous lineage of clonal workers that has infested commercial Scutellata colonies ever since (Kryger 2001; Baudry et al. 2004; Oldroyd et al. 2011). Over the past 23 years, the Clone has been responsible for what became known as the ‘Capensis Calamity’ (Allsopp 1992; Neumann and Moritz 2002). While new beekeeping practices have reduced rates of transmission, the Clone lineage remains highly virulent and is still responsible for the loss of hundreds of commercial Scutellata colonies each year (Cobey 1999).
The invasion of Scutellata colonies by the Clone appears to be largely dependent on apicultural practices (Moritz 2002; Neumann and Hepburn 2002; Dietemann et al. 2006a). Clone infestation is observed at only low levels in the wild Scutellata population, and only when the wild colonies are in contact with domestic
Figure 4 Map of South Africa (after Oldroyd et al. 2011), showing (I) the natural range of Capensis, (II) the stabile hybrid zone between Capensis and Scutellata and (III) the South African distribution of Scutellata, over much of which the Clone can now be found.
colonies (Härtel et al. 2006). Clones have diffi- culty invading Scutellata colonies without assis- tance (Moritz et al. 2008). However, once a Clone has successfully established in a host colony, the colony's downfall is all but inevitable.
When Clones enter a Scutellata colony, they activate their ovaries and produce queen-like mandibular gland secretions despite the presence of the host queen (Härtel et al. 2011). Clones thus establish themselves as pseudoqueens, and are tended to by host workers as if they were the rightful Scutellata queen of the colony (Figure 5). The host queen is soon lost as a result of lethal fighting (Moritz et al. 2003) and pheromonal competition (Dietemann et al. 2006b; Moritz et al. 2004). The presence of reproductively active pseudoqueens may suppress the development of later Clone offspring, resulting in the establish- ment of dominance hierarchies (Härtel et al. 2011), with only a small number of Clones reaching reproductive dominance within the host
colony (Martin et al. 2002). However, despite the suppression of reproduction in many Clone offspring, they rarely engage in work such as foraging or brood care (Martin et al. 2002).
Clone larvae manipulate host nurse workers, eliciting greater levels of feeding, with food that is more similar in composition to that of royal jelly, than the fare normally provided to mere workers (Calis et al. 2002). The resultant Clones have more queen-like characteristics than normal workers, including shorter developmental time, higher weight, larger spermatheca and larger number of ovarioles, while worker characteristics such as pollen combs and pollen baskets on their hind legs are suppressed (Wirtz and Beetsma 1972; Calis et al. 2002). Thus, the host colony is soon over run with Clone pseudoqueens and their offspring, which only adds to the burden of useless reproductive workers already afflicting the host colony. With time, the number of host workers dwindles and the colony inevitably
Figure 5 Reproductive Clones in a host Scutellata colony. i Darker-bodied Clones (circled in red) are tended to by host workers as if they were host queens. ii In the later stages of invasion, Clones lay dozens of eggs in host brood cells that should only hold one. Photos by B Oldroyd.
declines and collapses (Allsopp and Crewe 1993; Hepburn and Allsopp 1994).
2.3. A social cancer
The analogy of the honeybee colony as a ‘super organism’ is well established and compelling (e.g. Wheeler 1911; Seeley 1989; Moritz and
Southwick 1992; Moritz and Fuchs 1998; Amdam and Seehuus 2006; Hölldobler and Wilson 2008; Johnson and Linksvayer 2010; Seeley 2010; Page 2013). The queen can be compared to the gonads of a multicellular organ-
ism, supported by the somatic cells, a role played by workers. Somatic cells do not reproduce themselves, instead they make up the larger whole that enables the gametic cells (drones and virgin queens) to survive and propagate into the next generation. Multicellularity has been able to evolve because the cells that make up the multicellular organism are identical, having prop- agated from a single zygote. Similarly, worker bees forgo direct reproduction in favour of supporting the reproductive efforts of their queen, and through their work, allowing the colony to survive and send forth reproductive swarms. Like
the cells of a multicellular organism, the individ- uals of a honey bee colony are related and so kin selection theory provides an explanation for how individuals could evolve to sacrifice direct repro- duction in favour of propagating their genes through the reproductive success of related individuals (Hamilton 1964).
Cancer occurs in a multicellular organism when mutations in somatic cells result in cellular replication without restraint (Weinberg 1998). Similarly, in the honey bee colony, cheater workers regularly emerge that abandon reproductive self-restraint and reproduce at the expense of the colony (Barron et al. 2001; Beekman and Oldroyd 2008b; Châline et al. 2002; Holmes et al. 2013; Montague and Oldroyd 1998; Oldroyd et al. 1994). In the Capensis population, cheating occurs when daughters of the colony lay eggs in queen cells (Jordan et al. 2008; Allsopp et al. 2010; Holmes et al. 2010; Moritz et al. 2011). Thus, these workers can be compared to cancerous cells in a multicellular organism. This is taken a step further, when reproductive parasites invade non-natal colonies and begin competing over reproduction. We might view non-natal repro- ductive parasitism in the Capensis population as a kind of transmissible cancer. This is not without precedent in multicellular organisms. (See for example, the contagious facial tumours of the Tasmanian Devil Sarcophilus harrisii and the sexually transmitted cancer of domes- tic dog Canis lupus familiaris; Siddle and Kaufman 2013.)
The Clone is an extreme example of this phenomenon, a self-propagating ‘cancerous’ line- age that reproduces outside any constraint imposed by the colony, while taking full advantage of the resources it provides (Oldroyd 2002). The Clone goes further than most cancers of multicellular organisms, for it is sometimes able to survive the destruction of its host and transfer to another.
Moritz et al. (2008) regard the Clone as parasite with high virulence and low transmis- sibility, resulting from shortsighted within-host selection (Levin 1996). Under this model, the most virulent parasitic genotype outcompetes less virulent genotypes during the infection
phase of the invasion, before horizontal trans- mission occurs, resulting in a selection of a lineage with high virulence but low transmis- sion (Bull 1994).
As predicted by a scenario of ‘short-sighted evolution’, rates of horizontal Clone transmis- sion were not only undetectably small in a source-sink experimental setup without apicul- tural intervention, they were much lower than rates of transmission of Capensis workers taken from the endemic Capensis range (Moritz et al. 2008).
The Clone emerged after the movement of over 200 Capensis colonies into the Scutellata range (see above and Allsopp and Crewe 1993). Assuming that each colony comprised maybe 20,000 workers, made up of at least 20 patrilines (Palmer and Oldroyd 2000), the truckload of colonies comprised approximately 4 million worker genotypes and at least 4,000 patrilines. From these genotypes, a single Clonal lineage emerged, one selected for high virulence within Scutellata host colonies. Clonal reproduction then enabled this lineage to endure for generations, with its virulent genotype unchanged by sexual recombination.
2.4. Maintenance of heterozygosity in the Clone
Thelytoky in Capensis (automixis with central fusion, see above) carries the inherent feature of loss of heterozygosity. Specifically, wherever recombination exchanges genetic material be- tween chromosomes, there is a 1/3 chance that a locus that is heterozygous in the mother will become homozygous in offspring (Pearcy et al. 2006; Oldroyd et al. 2008; Engelstadter et al. 2010) (Figure 1). Therefore, ongoing generations of thelytoky should result in population-wide homozygosity at all loci that are free to recombine (Goudie et al. 2012b). Yet empirical studies have revealed levels of heterozygosity in the Clone that are remarkably high (Baudry et al. 2004; Neumann et al. 2011; Oldroyd et al. 2011).
Historically, the unexpectedly high levels of heterozygosity observed in the Clone were attrib- uted to a reduction in meiotic recombination
(Moritz and Haberl 1994; Baudry et al. 2004). However, Goudie et al. (2012a) demonstrate that a reduction in recombination is insufficient to ex- plain current levels of heterozygosity. Loss of heterozygosity in a thelytokous lineage is cumula- tive. Heterozygous mothers produce homozygous daughters at 1/3 the rate of recombination, while homozygous mothers produce homozygous off- spring exclusively (Engelstadter et al. 2010; Goudie et al. 2012b). Therefore, for any realistic level of recombination (whether reduced or not), homozygosity will inevitably accumulate. After 20 years of exclusive thelytokous reproduction, reduction in recombination cannot explain the maintenance of heterozygosity in the Clone at any but the most centromeric loci where recombi- nation is exceedingly rare (Goudie et al. 2012b).
Maintenance of heterozygosity in the Clone can instead be explained by selection against homozy- gous recombinants at genes that are subject to heterozygote advantage (Oldroyd et al. 2011; Goudie et al. 2012b). A key example of the effects of this kind of selection is the maintenance of heterozygosity at the complementary sex- determining locus (csd). Honey bees must be heterozygous at the csd for the female phenotype to be expressed, while haploid males are hemizy- gous. Homozygosity at the csd results in the production of a diploid male. Diploid males are inviable because they are eaten at early larval stages (Woyke 1963; Beye et al. 2003). Therefore, the csd locus is homozygous lethal, and any Clone offspring in which recombination results in homozygosity at this locus will be lost to cannibalism, thus main- taining heterozygosity at the csd in perpetuity.
The csd is a locus with known heterozygous advantage (overdominance). However, heterozy- gosity is observed throughout the Clone genome at loci that are unlinked to csd and presumed to be selectively neutral. How? There is now strong evidence that heterozygosity is maintained throughout much of the Clone's genome by selection acting on overdominant loci (Goudie et al. 2012b, 2014). These putative overdominant loci are theorised to be in linkage disequilibrium with the marker loci that are observed to be heterozy- gous. In support of this theory, the frequency of homozygosity is significantly higher in Clone eggs
than in it is in larvae and pupae, at both the csd and a range of neutral markers unlinked to the csd (Goudie et al. 2012b). These analyses show that recombination occurs at normal or near-normal rates in the Clone, resulting in the production of homozygotes, including diploid males. However, these recombinants are rapidly removed from the population, permanently retaining heterozygosity in the surviving Clones (Goudie et al 2012a, 2014). Goudie et al. (2014) mapped patterns of zygosity along chromosomes III and IV in the Clone to determine the evolutionary outcome of recombina- tion and selection. Loss of heterozygosity in a Clonal lineage is non-reversible, and a single recombination event will result in loss of heterozy- gosity at all markers located in a telomeric direction from the point of chiasmata, unless a second, concurrent recombination event results in restora- tion of heterozygosity (Figure 6). Yet in the Clone, complete loss of heterozygosity occurs in restricted regions, with subsequent restoration of heterozy- gosity in telomeric regions (Figure 7). This pattern of hetero/homo-zygosity along chromosomes sug- gests that overdominant genes located in the telomeric regions maintain heterozygosity, and indeed this pattern is observed at the csd on chromosome III. Goudie et al. (2014) therefore suggested that there are at least three overdominant genes maintaining heterozygosity on chromosome IV, and four genes (including the csd) that maintain
heterozygosity on chromosome III.
While low rates of recombination were once thought to maintain heterozygosity during thelytoky, growing evidence suggests that usu- ally high rates of recombination may in fact shape the evolution of the Capensis genome under thelytoky. The honey bee genome is characterised by very high rates of recombina- tion (4 times higher than most other taxa and 20 times higher than in humans) (Beye et al. 2006; Solignac et al. 2007). Furthermore, the honey bee has low levels of positive interference (Solignac et al. 2004), i.e. one recombination event does not suppress the probability of a second recombination event occurring nearby. Therefore, high rates of double recombination events within relatively small genetic distances are not unexpected in a honey bee genome. It is
Figure 6 i A single recombination event (orange X) will result in loss of heterozygosity at all telomeric markers (b, c, d). If a locus d (here linked to the marker c) is selectively overdominant, such a recombinant genotype will be selected against. ii Loss of heterozygosity at the marker B will only be observed if it is accompanied by a second, concurrent ‘rescue’ recombination event (blue X) which restores heterozygosity at locus D which is assumed to be under overdominant selection for heterozygosity.
these double-recombination events that are required to generate the genotypes that allow selection to maintain heterozygosity at isolated overdominant loci under selection, while het- erozygosity is lost along the majority of the chromosome (above, Goudie et al. 2013).
High rates of recombination and positive inter- ference in the honey bee has recently been linked to a high rate of gene conversion without crossover (non-crossover) events (Bessoltane et al. 2012). In fact, recombination events are more frequent in the honey bee genome than crossover events. Allelic gene conversion (the replacement of one allele with another at the same locus) can result in loss of heterozygosity during thelytoky. However, non- allelic gene conversion (the replacement of an allele with another from a different locus) could in fact increase genetic diversity, even in a clonal population. It is thus possible that gene conversion may counter loss of heterozygosity during thelytoky. The degree to which gene conversion occurs in Capensis remains to be established, as does the impact that gene conversion may have on the already-documented selective processes that retain heterozygosity in the Clone (Goudie et al. 2012b, 2013).
2.5. Clone drones
Recent evidence has shown that Clones do not reproduce exclusively via thelytoky, as had previ-
ously been assumed (Lattorff et al. 2005). Haploid male eggs were detected in the brood of Clone workers, at a frequency of one in eight (Goudie et al. 2012b). In larvae of the Clones, the frequency of haploid males dropped fivefold relative to eggs, suggesting strong selection against Clone males, though this selective removal may have arisen as a result of haploid male eggs being laid in worker cells. Preliminary evidence now suggests that a few Clone drones survive to maturity. A single adult haploid male carrying Clone alleles at all loci tested (n=9) was detected among 78 black drones collected from Clone-infested Scutellata colonies (Goudie et al., unpublished data). Thus, despite the apparent low frequency of adult Clone drones, our singular example (thus far) shows that some haploid males produced by the Clone lineage are able to reach maturity. It is therefore possible that Clone drones mate with Scutellata queens, resulting in introgression of Clone alleles in to the Scutellata population. This further raises the possibility of contagious parthenogenesis (Engelstadter et al. 2010) in which the rare production of males by clonal lineages leads to the transmission of alleles conferring asexuality into otherwise sexual populations.
2.6. The genetics of thelytoky in Capensis
Thelytoky in Capensis is thought to be con- trolled by a single recessive locus termed thelytoky
Figure 7 The pattern of zygosity along chromosomes III and IV, of the Clone incorporating a descriptive model for the maintenance of heterozygosity (Goudie et al 2013). Heterozygosity is maintained in blue regions by linkage to a heterozygosity-maintaining factor (HMF): the centromere, the csd, or putative genes under overdominant selection (a, b, c). As we move down the chromosome towards the telomere, heterozygosity is lost in purple regions as a result of a recombination event at points telomeric to a HMF, but restored by a second, concurrent recombination which occurred before the next HMF (e.g. heterozygosity is lost after the overdominant gene A on chromosome III, but restored before the gene B). In the yellow region of chromosome IV heterozygosity declines gradually, suggesting either incomplete selection, or the escape from selection by some sublineages due to double recombination events that were undetected in this analysis.
(th) (Lattorff et al. 2005). However, backcross experiments suggest that while th plays a major role in determining the thelytoky phenotype, the genetic basis of thelytoky may be a little more complex than is currently appreciated (Oldroyd et al., unpublished data). Furthermore, the frequent production of haploid eggs by Clones (with the putative genotype th,th) suggests that the th locus may not have complete expressivity (Goudie et al.
2012b). Alternatively, errors in thelytoky may be frequent in this lineage.
Under the single gene model for thelytoky, it has been proposed that differential splicing of the transcription factor CP2, a homolog for the Drosophila transcription factor gemini, results in the development of the thelytokous pheno- type (Jarosch et al. 2011). Splice forms of CP2 in Capensis are more similar to those of sexual
queens then arrhenotokous workers in other subspecies. Jarosch et al. (2011) suggest that thelytoky in Capensis may be determined by the lack of a short splice enhancer motif. Knockdown of this motif in arrhenotokous workers results in rapid ovary activation, which is one of a number of features that characterise the highly reproductive Capensis worker phe- notype.
/
2.7. Capensis and sex
Capensis, and in particular the Clone, pro- vides a valuable model with which to investi- gate the genetic and evolutionary consequences of thelytokous parthenogenesis, providing unique insights into the evolutionarily tradeoff between sex and asexuality that drives the distribution of reproductive strategies among animal taxa.
Sexual reproduction is the predominant form of reproduction among multicellular organisms (White 1984; Suomalainen et al. 1987), yet the near ubiquity of sexual reproduction remains an enduring evolutionary mystery. Many potential benefits of sex have been proposed and inves- tigated (see Otto and Gerstein 2006; Engelstadter 2008). These seek to deal with the fundamental question of how an allele imparting sexual reproduction could outcom- pete an allele causing asexual reproduction when sexuality reduces the reproductive poten- tial of a population by a factor of 2, as a consequence of the production of males that do not themselves reproduce (Maynard Smith 1978). In Capensis, we observe three unique female reproductive phenotypes, the queen, the worker and the Clone. Each of these utilise the same underlying genotype, however, the interplay of life history with the costs and benefits of sex and asexuality has resulted in the evolution of
distinctly different reproductive strategies.
2.8. The queen
At first, it appears perplexing that the Capensis queen forgoes thelytoky. Thelytokous reproduc- tion would allow a Capensis queen to produce
daughter queens that are related to her by unity (r
=1). Like certain thelytokous ant species, she could perhaps continue to employ sexual repro- duction to produce workers (Cataglyphis cursor, Pearcy et al. 2004; Wasmania auropunctata, Fournier et al. 2005; Vellonhovia emeryi, Kobayashi et al. 2008), gaining the best of both evolutionary worlds: effective genetic immortality in her reproductive offspring and genetic variabil- ity with its associated benefits (Jones et al. 2004; Mattila and Seeley 2007; Oldroyd and Fewell 2007; Seeley and Tarpy 2007) in her workers. Thelytoky is a very real evolutionary option for a Capensis queen. A virgin Capensis queen can reproduce both thelytokously and arrhenotokous when induced to start laying by double narcosis with CO2 (Allsopp and Crewe 1993; Oldroyd et al. 2008). Yet, despite the potential benefits, there is no evidence that mated Capensis queens ever lay thelytokous eggs (Jordan et al. 2008; Holmes et al. 2010; Moritz et al. 2011), providing strong evidence that for the Capensis queen, the costs of thelytoky outweigh the costs of sex.
A honey bee colony reproduces via the production of drones and swarms. The queen leaves the colony heading a swarm comprised of about half the workers, leaving behind a small number of queen cells containing her queen-destined daughters. One of these daugh- ters will inherit the original colony, while one or two others may head secondary swarm that has a lower survival than the first (Hepburn and Radloff 1998; Seeley 2010). Thus, queens trust their reproductive futures in a tiny number of daughter queens. Any reduction in larval via- bility associated with thelytoky may therefore have a substantial impact on a queen's fitness (her larva may be usurped by that of a worker), which is compounded by the absence of the many benefits associated with sexual reproduc- tion (Otto and Gerstein 2006).
A queen that produces daughter queens asexu- ally and daughter workers sexually would come into direct conflict with her worker daughters. She would share twice as many alleles with her own thelytokously-produced queen daughter (r=1) than she would with the thelytokously-produced daugh- ters of her sexually produced worker (r=0.5;
Figure 8). A queen that produces both worker and queen offspring thelytokously eliminates this com- petition, but in the process, massively reduces the genetic diversity of her work force and so potentially the fitness of the colony she relies on to raise her reproductive offspring (Figure 8). However, a sexual queen is equally related to her sexual daughter as she is to her thelytokously- produced granddaughter (Figure 8). Therefore, provided her workers are working (and not just breeding, Hillesheim et al. 1989), a queen is predicted to be indifferent to the production of new queens by natal workers (Greeff 1996; Beekman and Oldroyd 2008a).
2.9. The typical Capensis worker
Thelytoky massively increases the reproduc- tive potential of a Capensis worker, enabling her to produce diploid daughters and to compete with her mother and fellow workers over the production of new queens. Not only does thelytoky increase the reproductive potential of the Capensis worker, it fundamentally alters the kin structure of a Capensis colony relative to that of an arrhenotokous colony, eliminating, or greatly reducing, the selective pressures that normally drive workers to suppress the repro- ductive proclivities of their worker sisters (Greeff and Villet 1993; Moritz et al. 1999). Honey bee queens are extremely polyandrous (Palmer and Oldroyd 2000) and as a result workers within a colony are mainly half sisters. Thus, in an arrhenotokous colony, a worker is more closely related to the sons of her mother (r
=0.25) than to the sons of a fellow worker (r= 0.125). While a worker might benefit from producing her own sons (r=0.5), collectively workers prefer to raise the sons of their mother (Ratnieks 1988). As a result, worker policing has evolved, where workers eat eggs that have not been laid by the queen (Ratnieks and Visscher 1989). In contrast, Capensis workers can benefit immensely from personal reproduc- tion, while the queen and other workers are largely indifferent to it, provided it does not unduly reduce colony productivity (Beekman et al. 2002, 2009; Greeff and Villet 1993; Moritz
et al. 1999; Pirk et al. 2002). While the Capensis worker still benefits more from pro- ducing her own offspring than raising the offspring of another, she is indifferent to whether the offspring of other females are produced by workers or queens. Thus, instead of policing, directed worker competition is expected to evolve, and is observed (Jordan et al. 2008; Moritz et al. 1996, 2011).
As described above, thelytoky incurs a cost in Capensis workers; a 1/3 loss of heterozygos- ity per generation for any locus that is free to recombine. So, for example, 1/3 of eggs laid by Capensis workers should be inviable diploid males. However, the reproductive Capensis worker takes advantage of reproductive oppor- tunities that are otherwise unavailable. Unlike the Capensis queen, sex is not an evolutionary option for the worker, while thelytoky provides a worker with a window of opportunity to be genetically reincarnated to the queen phenotype. A worker's thelytokously-produced daughter queen subsequently reproduces sexually, and so the cost of thelytoky in the worker is only paid over a single generation; loss of heterozy- gosity does not compound once the worker is reincarnated as a queen.
The thelytokous worker has everything to gain and little to lose though thelytokous parthenogenesis, particularly when frequency- dependent selective forces maintain ‘cheater’ parasitic lineages at low levels that do not jeopardise the stability of the eusocial colony (Hillesheim et al. 1989).
2.10. The Clone
The introduction of highly reproductive Capensis workers to Scutellata colonies enables the emergence of asexual lineages that are completely liberated from reliance on a sexual queen for their vicarious reproduction. However, for thelytoky to endure over evolutionary time, a cost must be paid. Maintenance of heterozygosity by selection requires the removal of homozygous recombinant offspring each and every generation. To be specific, heterozygosity will be maintained at a locus provided that the number of homozygotes
a b c
0.5
Figure 8 The three evolutionary options for Capensis queens. The queen in each pedigree is circled in red, all values refer to her relatedness (r) to each individual. a A queen that produces workers sexually (ii) and new queens thelytokously (ii) maximises her relatedness to her queen daughters while maintaining genetic diversity in her worker force. However, she will come into conflict with her daughters. She is twice as related to her own queen daughter (ii) as she is to a worker-produced queen (iii). b A queen that produces both worker (i) and queen (ii) daughters thelytokously avoids competition. She is equally and maximally related to her own queen daughter (ii) as she is to a worker-produced queen (iii). However, she has much reduced genetic diversity in her colony, which may suffer from reduced disease resistance and less efficient task allocation. c A queen that produces both worker (i) and queen (ii) daughters sexually also avoids conflict with her female offspring, while maintaining genetic diversity in the colony In this scenario, which is what we also see in reality, the queen only shares half her alleles with her queen daughters. However, when we take into account the evolutionary alternatives (a and b), we see that this strategy maximises queen fitness.
being removed by selection is equal to or greater than the number being produced by recombination (Goudie et al. 2012b). Therefore, for a thelytokous lineage such as the Clone to endure, the benefits of thelytoky must outweigh the per-generational cost in reduced viability, which is necessary to maintain the integrity of the clonal genome. We (Goudie et al. 2012b) proposed that the parasitic life history of the Clone does indeed make it ideally suited to enduring this cost.
During an invasion, Clone workers lay a massive number of eggs. Eventually, brood cells that should only hold one egg become packed with dozens of Clone progeny (Figure 5). Only a tiny proportion of these eggs can ever be expected to hatch, let alone emerge as an adult, survive colony collapse and continue the invasive cycle (Martin et al. 2002; Neumann and Hepburn 2002). And for a Clone, the production of these eggs is cheap; she is waited on, wing and tarsis, by her hosts, taking no part in non-reproductive tasks, such as foraging and brood care. Any of her offspring that emerge
are abandoned to the care of their hosts, from whom Clones elicit level of attention normally reserved for royalty (Beekman et al. 2000; Allsopp et al. 2003). A Clone can therefore dedicate her life to producing thelytokous eggs, in the hope that some will reach maturity. High rates of reproduc- tion, low maternal investment and concordantly high mortality are an inherent part of the Clone's parasitic reproductive strategy—when the vast majority of eggs cannot be raised to maturity, it hardly matters if many of them are inviable.
2.11. Of queens, workers and Clones
While thelytoky imposes high costs, it allows the Clone to exploit a new niche that would otherwise be unavailable, that of social parasitism. Parasitism is both the means by which the Clone benefits from going without sex, and the means in which it is able to endure the costs of thelytoky. Workers from the sexual Capensis population, in contrast, play the odds, giving thelytoky a go
because they have no other avenue for direct reproduction, while still being relatively assured of the indirect benefits of a eusocial existence. In the queen, we see the more standard outcome to the evolutionary tradeoff between sex and asexu- ality, the costs of thelytoky may be too high a price when the queen's reproductive future, and that of the colony, is vested in a small number of potentially reproductive daughters.
Using Capensis and its Clone as a model, we suggest that the costs and benefits of sex and asexuality should be considered in a more conditionally than is often the case. The specific life history of a population, and the outcomes of the mode of thelytoky it employs, must be examined to account for where costs and benefits are imposed, and where they can be endured.
3. THE EVOLUTION OF THELYTOKY IN APIS
3.1. Thelytoky in Apis more broadly
Capensis appears to be the only honey bee in which thelytoky is ubiquitous. However, Mackensen (1943) reported that approximately one percent of eggs produced by virgin queens of the Italian (Apis mellifera ligustica) and Caucasian (Apis mellifera caucasica) subspecies were female, the result of thelytokous partheno- genesis. (Mackensen's experimental queens had been exposed to double CO2 narcosis, which induces oviposition in honey bee queens, normal- ly resulting in the production of arrhenotokous males.) The low frequency of thelytokous repro- duction may well be the result of errors in arrhenotokous parthenogenesis. However, the regularity with which thelytokous offspring was observed by Mackensen (1943) suggests that thelytoky is a threshold character that can be released with relatively small genomic, and perhaps environmental, changes.
This conjecture is supported by the frequency with which thelytokous reproductive systems are being identified in another taxa of eusocial Hymenoptera. Thelytoky is relatively common in ants (Rabeling and Kronauer 2013) and the number of known thelytokous ant species has
dramatically increased over the last few years, as more species are investigated with molecular techniques. Importantly, thelytoky in ants ap- pears to be associated with invasive life histo- ries (Rabeling and Kronauer 2013).
While not a honey bee, the solitary little carpenter bee Ceratina dellatorreana has been reported to reproduce thelytokously. As with many ants, thelytoky in the C. dellatorreana was observed in an invasive population, where it is hypothesised to have facilitated its intro- duction (Daly 1966). However, no further information is available on C. dellatorreana.
It appears very possible that more thelytokous bees are waiting to be discovered. Even well- studied populations in which both males and females are present can be producing thelytokous females that are not detectable until we look for them explicitly, usually with molecular techniques.
3.2. Thelytoky in A. cerana
Evidence is currently accumulating that indicates that another species of honey bee, the Asiatic honey bee A. cerana, reproduces thelytokously (Holmes, unpublished data).
In recent years, an invasive population of A. cerana was identified in Queensland, Australia (Koetz 2013). This population was probably founded by a single reproductive swarm, and as such the population has limited genetic diversity. We hypothesise that thelytoky may have enabled the successful establishment of this invasive population. A disturbing possibility is that inter- specific matings between A. cerana males and A. mellifera queens may induce thelytoky in A. mellifera queens (unpublished data).
3.3. The evolution of thelytoky in Capensis
Thelytoky may have evolved to be ubiquitous in Capensis during periods of the Pleistocene in which rising sea levels isolated the Cape Peninsula from the rest of the African continent (Ruttner 1977). It was once feared that the world's only known thelytokous bee would be overrun by the more aggressive and widespread Scutellata subspecies (a.k.a. the African killer bee; Ruttner
1977). This fear is ironic in hindsight, given that we now know that Scutellata comes off second best when bought into contact with Capensis. In reality, a stable hybrid zone that neither subspecies is able to cross without human intervention (Beekman et al. 2008) now separates the two populations. Hybrid or mixed colonies of Capensis and Scutellata are assumed to suffer from reduced fitness (Beekman et al. 2008, 2012), though evidence for this hypothesis is currently lacking. Scutellata drones and virgin queens may outcompete Capensis at mating leks. However, even low frequencies of Capensis genotypes within a mixed subspecies colony is expected to result in a misallocation of resources by easily duped Scutellata workers, leading to the produc- tion of more reproductive Capensis workers, and a breakdown in regulation of worker reproduction (Beekman et al. 2008).
It has been hypothesised that thelytoky in Capensis originally evolved in response to high rates of queen loss on the windy and often inclement Cape Peninsular (Tribe 1983). Thelytoky does indeed provide Capensis with the means to produce a new queen when arrhenotokous subspecies might fail. However, given the global range of the honey bee, it seems highly unlikely that any environmental conditions experienced by Capensis are so unique that they alone have driven the evolution of such a distinctly divergent reproductive strategy.
We suggest that the evolution of thelytoky in Capensis was facilitated by genetic drift in a small, isolated population. While queen replace- ment may have played a role in selecting for thelytoky, a range of other factors would have been required for thelytoky to become ubiqui- tous in the population. Even assuming that thelytoky occurs at low frequency in otherwise arrhenotokous honey bee populations, workers laying thelytokous eggs in drone cells run into a genetic dead end, because these eggs will never develop to queens. Thus, thelytoky would not be selected for until such time as it co-occurs with a heritable behavioural variant in which workers target their reproductive efforts to queen cells. Furthermore, thelytoky emerging from a background of worker arrhenotoky does
so in an environment of intense worker policing (Ratnieks 1988). Selection to reduce worker policing will not occur until after thelytoky has become ubiquitous. Thus, policing acts as an evolutionary barrier, reducing or eliminating any immediate payoff from thelytoky.
If thelytoky is to reach a high frequency in a honey bee population, multiple factors must fall into place concurrently: a thelytokous mode of worker reproduction, targeting of worker repro- duction to queen cells, and the relaxation of worker policing. In a large, outbred population the suppression of worker reproduction by worker policing may significantly reduce variance in both the mode and target of worker's reproductive efforts. Thus, thelytoky is unlikely to emerge. However, in a small, isolated population, faced with additional pressures such as a high rate of queen loss (above), genetic drift and founder effects may have resulted in the necessary combination of factors falling into place for thelytoky to reach a stable point in the population.
4. CONCLUSION
The honey bee has played an important role in driving and informing evolutionary theory, a role that shows no sign of ending soon. Here, we have shown that the honey bee, and particularly the Capensis subspecies, has much to contribute to questions concerning the evolution of sex and asexuality. In Capensis, sex and asex co-exist, distributed among castes and lineages that share the same genetic background. Differing life histories results in divergent outcomes when the costs and benefits of sex and asexuality come into conflict. We suggest that the broader question of why sex evolved from ancestral asex, and how it has been maintained, should be addressed with an eye for more conditional costs and benefits.
We further propose that there may be broader implications to the insights provided by bees, ants and the haplodiploid Hymenoptera in general. In these species, a form of asexual reproduction, arrhenotoky, is ancestral. While arrhenotokous species still require sex for the production of females, this reproductive system is hypothesised to predispose haplodiploids to the evolution of true,
thelytokous, asexuality (Engelstadter 2008; Rabeling and Kronauer 2013). In non- arrhenotokous species, the transition from sex back to asexuality is not as easy (Engelstadter 2008) and so the evolution of sex may be, in most circum- stances, a one-way street. The question of “why is sex always better then asex?” then becomes “why is sex ever better then asex?” Sexual reproduction may evolve in a species during a period in which environmental conditions are such that the evolu- tionary tradeoff between sex and asex is similar to that faced by the Capensis queen, i.e. the need to invest maximally in a limited number of offspring. However, having taken this route, they cannot simply switch back to asex when conditions change. And so it is possible that there are many potential Clones waiting to emerge, but for these species asexuality is not a realistic evolutionary option, despite the benefits it may confer.
Parthénogenèse thélytoque chez l'abeille
Apis mellifera / Apis mellifera capensis / reproduction asexuée / thélytocie / parasitisme reproducteur
Thelytökie bei Honigbienen
Apis mellifera / Apis mellifera capensis / asexuale Reproduction / Thelytökie / reproduktiver Parasitismus
REFERENCES
Allsopp, M. (1992) The Capensis calamity. South African Bee Journal 64, 52–55
Allsopp, M., Crewe, R.M. (1993) The Cape honeybee as a Trojan horse rather than the hordes of Jenghiz Khan. Am. Bee. J. 133, 121–123
Allsopp, M., Calis, J.M., Boot, W. (2003) Differential feeding of worker larvae affects caste characters in the Cape honeybee, Apis mellifera capensis. Behav. Ecol. Sociobiol. 54, 555–561
Allsopp, M., Beekman, M., Gloag, R., Oldroyd, B.P. (2010) Maternity of replacement queens in the thelytokous Cape honey bee Apis mellifera capensis. Behav. Ecol. Sociobiol. 64, 567–574
Amdam, G.V., Seehuus, S.C. (2006) Order, disorder, death: lessons from a superorganism. Adv. Cancer Res. 95, 31–60
Amdam, G.V., Norberg, K., Fondrk, M.K., Page, R.E. (2004) Reproductive ground plan may mediate colony-level selection effects on individual foraging behavior in honey bees. Proc. Natl. Acad. Sci. USA 101, 11350–11355
Barron, A.B., Oldroyd, B.P., Ratnieks, F.L.W. (2001) Worker reproduction in honey-bees (Apis) and the anarchic syndrome: a review. Behav. Ecol. Sociobiol. 50, 199–208
Baudry, E., Kryger, P., Allsopp, M., Koeniger, N., Vautrin, D., et al. (2004) Whole-genome scan in thelytokous-laying workers of the Cape honeybee (Apis mellifera capensis): central fusion, reduced recombination rates and centromere mapping using half-tetrad analysis. Genetics 167, 243–252
Beekman, M., Oldroyd, B. (2008a) Who is the Queen's mother?
Royal cheats in social insects. J. Biosci. 33, 159–161
Beekman, M., Oldroyd, B.P. (2008b) When workers disunite: intraspecific parasitism in eusocial bees. Annu. Rev. Entomol. 53, 19–37
Beekman, M., Calis, J.N.M., Boot, W.J. (2000) Parasitic honeybees get royal treatment. Nature 404, 723
Beekman, M., Good, G., Allsopp, M., Radloff, S., Pirk, C., et al. (2002) A non-policing honey bee colony (Apis mellifera capensis). Naturwissenschaften 89, 479–482
Beekman, M., Allsopp, M.H., Wossler, T.C., Oldroyd,
B.P. (2008) Factors affecting the dynamics of the honeybee (Apis mellifera) hybrid zone of South Africa. Heredity 100, 13–18
Beekman, M., Allsopp, M.H., Jordan, L.A., Lim, J., Oldroyd, B.P. (2009) A quantitative study of worker reproduction in queenright colonies of the Cape honey bee, Apis mellifera capensis. Mol. Ecol. 18, 2722–2727
Beekman, M., Allsopp, M.H., Lim, J., Goudie, F., Oldroyd, B.P. (2011) Asexually produced Cape honeybee queens (Apis mellifera capensis) repro- duce sexually. J. Hered. 102, 562–566
Beekman, M., Allsopp, M., Holmes, M., Lim, J., Noach- Pienaar, L.-A., et al. (2012) Racial mixing in South African honeybees: the effects of genotype mixing on reproductive traits of workers. Behav. Ecol. Sociobiol. 66, 897–904
Bessoltane, N., Toffano-Nioche, C., Solignac, M., Mougel, F. (2012) Fine scale analysis of crossover and non-crossover and detection of recombination sequence motifs in the honeybee (Apis mellifera). PLoS Biol. 7, e36229
Beye, M., Hasselmann, M., Fondrk, M.K., Page, R.E., Omholt, S.W. (2003) The gene cds is the primary signal for sexual development in the honeybee and encodes an SR-type protein. Cell 114, 419–429
Beye, M., Gattermeier, I., Hasselmann, M., Gemp, T., Schioett, M., et al. (2006) Exceptionally high levels of recombination across the honey bee genome. Genome Res. 16, 1339–1344
Blacher P., Yagound B., Lecoutey E., Devinne P., Chameron S., et al. (2013) Drifting behaviour as
an alternative reproductive strategy for social insect workers. Proc. R. Soc. B. doi: http://dx.doi.org/ 10.1098/rspb
Bull, J.J. (1994) Perspective—virulence. Evolution 48,
1423–1437
Calis, J.N.M., Boot, W.J., Allsopp, M.H., Beekman, M. (2002) Getting more than a fair share: nutrition of worker larvae related to social parasitism in the Cape honey bee Apis mellifera capensis. Apidologie 33, 193–202
Châline, N., Ratnieks, F.L.W., Burke, T. (2002) Anarchy in the UK: detailed genetic analysis of worker reproduction in a naturally occurring British anar- chistic honeybee, Apis mellifera, colony using DNA microsatellites. Mol. Ecol. 11, 1795–1803
Chapman, N.C., Nanork, P. , G loag, R.S., Wattanachaiyingcharoen, W., Beekman, M., et al. (2009) Queenless colonies of the Asian red dwarf honey bee (Apis florea) are infiltrated by workers from other queenless colonies. Behav. Ecol. 20, 817–820
Cobey, S. (1999) The African bee, Apis mellifera scutellata, threatened in her South African homeland by the Cape bee, Apis melliera capensis. Am. Bee. J. 139, 462–467
Daly, H.V. (1966) Biological studies on Ceratina dallatorreana, an alien bee in California which reproduces by parthenogenesis (Hymenoptera: Apoidea). Ann. Entomol. Sco. Am. 59, 1138–1154
Dietemann, V., Lubbe, A., Crewe, R.M. (2006a) Human factors facilitating the spread of a parasitic honey bee in South Africa. J. Econ. Entomol. 99, 7–13
Dietemann, V., Pflugfelder, J., Härtel, S., Neumann, P., Crewe, R. (2006b) Social parasitism by honeybee workers (Apis mellifera capensis Esch.): evidence for pheromonal resistance to host queen's signals. Behav. Ecol. Sociobiol. 60, 785–793
Engelstadter, J. (2008) Constraints on the evolution of asexual reproduction. BioEssays 30, 1138–1150
Engelstadter, J., Sandrock, C., Vorburger, C. (2010) Contagious parthenogenesis, automixis, and a sex determination meltdown. Evolution 65, 501–511
Fournier, D., Estoup, A., Orivel, R.M., Foucaud, J., Jourdan, H., et al. (2005) Clonal reproduction by males and female in the little fire ant. Nature 435, 1230–1234
Gladyshev, E., Meselson, M. (2008) Extreme resistance of bdelloid rotifers to ionizing radiation. Proc. Natl. Acad. Sci. USA 105, 5139–5144
Goudie, F., Allsopp, M.H., Beekman, M., Lim, J., Oldroyd, B.P. (2012a) Heritability of worker ovar- iole number in the Cape honey bee Apis mellifera capensis. Insectes Soc. 59, 351–359
Goudie, F., Allsopp, M.H., Beekman, M., Oxley, P.R., Lim, J., et al. (2012b) Maintenance and loss of heterozy- gosity in a thelytokous lineage of honey bees (Apis mellifera capensis). Evolution 66, 1897–1906
Goudie, F., Allsopp, M.H., Oldroyd, B.P. (2014) Selection on overdominant genes maintains het- erozygosity along multiple chromosomes in a clonal lineage of honey bee. Evolution 68, 125– 136
Greeff, J.M. (1996) Effects of thelytokous worker reproduction on kin-selection and conflict in the Cape honeybee, Apis mellifera capensis. Philos. Trans.: Biol Sci. 351, 617–625
Greeff, J.M., Villet, M.H. (1993) Deducing the coeffi- cient of relationship by the amount of recombination produced during automictic parthneogenesis. Heredity 70, 499–502
Gruber M., Hoffmann B., Ritchie P., Lester P.. (2010) Crazy ant sex: genetic caste determination, clonality, and inbreeding in a apopulation of invasive Yellow crazy ants. In: Nash D.R., den Boer S.P.A., Fine Licht H.H., and Boomsma J.J. (Eds.), XVI Congress of the International Union for the Study of Social Insects, Copenhagen, Denmark.
Hagimori, T., Abe, Y., Date, S., Miura, K. (2006) The first finding of a Rickettsia bacterium associated with parthenogenesis induction among insects. Curr. Microbiol. 52, 97–101
Hamilton, W.D. (1964) The genetical evolution of social behaviour. I & II. J. Theor. Biol. 7, 1–52
Härtel, S., Neumann, P., Kryger, P., von der Heide, C., Moltzer, G., et al. (2006) Infestation levels of Apis mellifera scutellata swarms by socially parasitic Cape honeybee workers (Apis mellifera capensis). Apidologie 37, 462–470
Härtel, S., Wossler, T., Moltzer, G.-J., Crewe, R., Moritz, R.A., et al. (2011) Pheromone-mediated reproduc- tive dominance hierarchies among pseudo-clonal honeybee workers (Apis mellifera capensis). Apidologie 42, 659–668
Hepburn, H.R., Allsopp, M.H. (1994) Reproductive conflict between honeybees: usurpation of Apis mellifera scutellata colonies by Apis mellifera capensis. Suid-Afrikaanse Tydskrif vir Wetenskap 90, 247–249
Hepburn, H.R., Crewe, R.M. (1990) Defining the Cape honeybee: reproductive traits of queenless workers. S. Afr. J. Sci. 86, 524–527
Hepburn, H.R., Crewe, R.M. (1991) Portrait of the Cape honeybee, Apis mellifera capensis. Apidologie 22, 567–580
Hepburn, H.R., Radloff, S.E. (1998) Honeybees of Africa. Springer, Berlin
Heubel, K.U., Rankin, D.J., Kokko, H. (2009) How to go extinct by mating too much: population consequences of male mate choice and efficiency in a sexual-asexual species complex. Oikos 118, 513–520
Hillesheim, E., Koeniger, N., Moritz, R.F.A. (1989) Colony performance in honeybees (Apis mellifera capensis Esch.) depends on the proportion of subordinate and dominant workers. Behav Ecol Sociobiol 24, 291–296
Hölldobler, B., Wilson, E.O. (2008) The superorganism: the beauty, elegance, and strangeness of insect societies. W. W Norton, New York
Holmes, M.J., Oldroyd, B.P., Allsopp, M.H., Lim, J., Wossler, T.C., et al. (2010) Maternity of emergency queens in the Cape honey bee, Apis mellifera capensis. Mol. Ecol. 19, 2792–2799
Holmes, M.J., Oldroyd, B.P., Duncan, M., Allsopp, M.H., Beekman, M. (2013) Cheaters sometimes prosper: targeted worker reproduction in honeybee (Apis mellifera) colonies during swarming. Mol. Ecol. 22, 4298–4306
Huigens, M.E., Stouthamer, R. (2003) Parthenogenesis associated with Wolbachia. In: Bourtzis, K., Miller,
T.A. (eds.) Insect symbiosis, pp. 247–266. CRC Press, Boca Raton
Jarosch, A., Stolle, E., Crewe, R.M., Moritz, R.F.A. (2011) Alternative splicing of a single transcription factor drives selfish reproductive behavior in hon- eybee workers (Apis mellifera). Proc. Natl. Acad. Sci. USA 108, 15282–15287
Johnson, B.R., Linksvayer, T.A. (2010) Deconstructing the superorganism: social physiology, reproductive groundplans, and sociogenomics. Q. Rev. Biol. 85, 57–79
Jones, J., Myerscough, M., Graham, S., Oldroyd, B.P. (2004) Honey bee nest thermoregulation: diversity promotes stability. Science 305, 402–404
Jordan, L.A., Allsopp, M.H., Oldroyd, B.P., Wossler, T.C., Beekman, M. (2008) Cheating honeybee workers produce royal offspring. Proc. R. Soc. Lond. B. Biol. Sci. 275, 345–351
Keller, L. (2007) Uncovering the biodiversity of genetic and reproductive systems: time for a more open approach—American Society of Naturalists E.O. Wilson award winner address. Am. Nat. 169, 1–8
Kobayashi, K., Hasegawa, E., Ohkawara, K. (2008) Clonal reproduction by males of the ant Vollenhovia emeryi (Wheeler). Entomol. Sci. 11, 167–172
Koetz A. (2013) The Asian honey bee (Apis cerana) and its strains—with special focus on Apis cerana Java genotype. Literature review. Brisbane.
Kryger, K. (2001) The Capensis pseudo-clone, a social parasite of African honey bees. In: Menzel, R., Rademacher, E. (eds.) International Union for the Study of Social Insects, p. 208. IUSSI, Berlin
Lattorff, H.M.G., Kryger, P., Moritz, R.F.A. (2005) Queen developmental time and fitness consequences for queens of clonal social parasitic honeybees (A. m. capensis) and its host A. m. scutellata. Insectes Soc 52, 238–241
Levin, B.R. (1996) The evolution and maintenance of virulence in microparasites. Emerg. Infect. Dis. J 2, 93–102
Mackensen, O. (1943) The occurrence of parthenoge- netic females in some strains of honey-bees. J. Econ. Entomol. 36, 465–467
Maynard Smith, J. (1978) The Evolution of Sex.
Cambridge University Press
Mark Welch, J.L., Mark Welch, D.B., Meselson, M. (2004) Cytogenetic evidence for asexual evolution of bdelloid rotifers. Proc. Natl. Acad. Sci. USA 101, 1618–1621
Martin, S., Wossler, T., Kryger, P. (2002) Usurpation of African Apis mellifera scutellata colonies by para- sitic Apis mellifera capensis workers. Apidologie 33, 215–232
Mattila, H.R., Seeley, T.D. (2007) Genetic diversity in honey bee colonies enhances productivity and fitness. Science 317, 362–364
Montague, C.E., Oldroyd, B.P. (1998) The evolution of worker sterility in honey bees: an investigation into a behavioral mutant causing failure of worker policing. Evolution 52, 1408–1415
Moritz, R. (2002) Population dynamics of the Cape bee phenomenon: the impact of parasitic laying worker clones in apiaries and natural populations. Apidologie 33, 233–244
Moritz, R.F.A., Fuchs, S. (1998) Organization of honeybeee colonies: characteristics and conse- quences of a superorganism concept. Apidologie 29, 7–21
Moritz, R.F., Haberl, M. (1994) Lack of meiotic recombination in thelytokous parthenogenesis of laying workers of Apis mellifera capensis (the Cape honeybee). Heredity 73, 98–102
Moritz, R.F.A., Southwick, E.E. (1992) Bees as super- organisms. Springer, Berlin
Moritz, R.F.A., Kryger, P., Allsopp, M.H. (1996) Competition for royalty in bees. Nature 384, 31
Moritz, R.F.A., Kryger, P., Allsopp, M.H. (1999) Lake of worker policing in the Cape honeybee (Apis mellifera capensis). Behaviour 136, 1079–1092
Moritz, R.F.A., Pflugfelder, J., Crewe, R.M. (2003) Lethal fighting between honeybee queens and parasitic workers (Apis mellifera). Naturwissenschaften 90, 378–381
Moritz, R.F.A., Lattorff, H.M.G., Crewe, R.M. (2004) Honeybee workers (Apis mellifera capensis) compete for producing queen-like pheromone signals. Proc. R. Soc. Lond. B. Biol. Sci. 271(Supplement 3), S98– S100
Moritz, R., Pirk, C.W.W., Hepburn, H.R., Neumann, P. (2008) Short-sighted evolution of virulence in parasitic honeybee workers (Apis mellifera capensis Esch.). Naturwissenschaften 95, 507–513
Moritz, R.F.A., Lattorff, H.M.G., Crous, K.L., Hepburn,
H.R. (2011) Social parasitism of queens and workers in the Cape honeybee (Apis mellifera capensis). Behav. Ecol. Sociobiol. 65, 735–740
Nanork, P., Paar, J., Chapman, N.C., Wongsiri, S., Oldroyd, B.P. (2005) Asian honey bees parasitize the future dead. Nature 437, 829
Nanork, P., Chapman, N.C., Wongsiri, S., Lim, J., Gloag, R.S., et al. (2007) Social parasitism by workers in queenless and queenright Apis cerana colonies. Mol. Ecol. 16, 1107–1114
Neumann, P., Hepburn, H.R. (2002) Behavioural basis for social parasitism of Cape honeybees (Apis mellifera capensis). Apidologie 33, 165–192
Neumann, P., Moritz, R.F.A. (2002) The Cape honeybee phenomenon: the sympatric evolution of a social parasite in real time? Behav. Ecol. Sociobiol. 52, 271–281
Neumann, P., Radloff, S.E., Moritz, R.F.A., Hepburn, H.R., Reece, S.L. (2001) Social parasitism by honeybee workers (Apis mellifera capensis Escholtz): host finding and resistance of hybrid host colonies. Behav. Ecol. 12, 419–428
Neumann, P., Härtel, S., Kryger, P., Crewe, R.M., Moritz, R.F.A. (2011) Reproductive division of labour and thelytoky result in sympatric barriers to gene flow in honeybees (Apis mellifera L.). J. Evol. Biol. 24, 286–294
Normark, B.B. (2003) The evolution of alternative genetic systems in insects. Annu. Rev. Entomol. 48, 397–423
Oldroyd, B.P. (2002) The Cape honeybee: an example of a social cancer. Trends Ecol. Evol. 17, 249–251
Oldroyd, B.P., Beekman, M. (2008) Effects of selection for honey bee worker reproduction on foraging traits. PLoS Biol. 6, e56
Oldroyd, B.P., Fewell, J.H. (2007) Genetic diversity promotes homeostasis in insect colonies. Trends Ecol. Evol. 22, 408–413
Oldroyd, B. P., Smolenski, A. J., Cornuet, J.-M., Crozier,
R. H. (1994) Anarchy in the beehive. Nature 371,
749
Oldroyd, B.P., Allsopp, M.H., Gloag, R.S., Lim, J., Jordan, L.A., et al. (2008) Thelytokous partheno- genesis in unmated queen honeybees (Apis mellifera capensis): central fusion and high recombination rates. Genetics 180, 359–366
Oldroyd, B.P., Allsopp, M.H., Lim, J., Beekman, M. (2011) A thelytokous lineage of socially parasitic honey bees has retained heterozygosity despite at least 10 years of inbreeding. Evolution 65, 860–868
Onions, G.W. (1912) South African ‘fertile worker bees’.
Agr. J. U. S. Afr. 1, 720–728
Otto, S.P., Gerstein, A.C. (2006) Why have sex? The population genetics of sex and recombination. Biochem. Soc. Trans. 34(Part 4), 519–522
Page, R.E. (2013) The spirit of the hive. The mecha- nisms of social evolution. Harvard University Press, Cambridge, MA
Palmer, K.A., Oldroyd, B.P. (2000) Evolution of multiple mating in the genus Apis. Apidologie 31, 235–248
Pearcy, M., Aron, S., Doums, C., Keller, L. (2004) Conditional use of sex and parthenogenesis for worker and queen production in ants. Science 306, 1780–1783
Pearcy, M., Hardy, O., Aron, S. (2006) Thelytokous parthenogenesis and its consequences on inbreeding in an ant. Heredity 96, 377–382
Phiancharoen, M., Pirk, C.W.W., Radloff, S., Hepburn,
R. (2010) Clinal nature of the frequencies of ovarioles and spermathecae in Cape worker honey- bees, Apis mellifera capensis. Apidologie 41, 129– 134
Pirk, C.W.W., Neumann, P., Hepburn, H.R. (2002) Egg laying and egg removal by workers are positively correlated in queenright Cape honeybee colonies (Apis mellifera capensis). Apidologie 33, 203–211
Rabeling, C., Kronauer, D.J.C. (2013) Thelytokous parthenogenesis in eusocial hymenoptera. Annu. Rev. Entomol. 58, 273–292
Ratnieks, F.L.W. (1988) Reproductive harmony via mutual policing by workers in eusocial Hymenoptera. Am. Nat. 132, 217–236
Ratnieks, F.L.W., Visscher, P.K. (1989) Worker policing in honeybees. Nature 342, 796–797
Ravary, F., Jaisson, P. (2004) Absence of individual sterility in thelytokous colonies of the ant Cerapachys biroi Forel (Formicidae, Cerapachyinae). Insectes Soc. 51, 67–73
Ruttner, F. (1977) The problem of the cape bee (Apis mellifera capensis Escholtz): parthenogenesis—size of population—evolution. Apidologie 8, 281–294
Seeley, T.D. (1989) The honey bee colony as a superorganism. Am. Scientist 77, 546–553
Seeley, T.D. (2010) Honeybee democracy. Princeton University Press, Princeton, NJ
Seeley, T.D., Tarpy, D.R. (2007) Queen promiscuity lowers disease within honeybee colonies. Proc. R. Soc. B 274, 67–72
Siddle, H.V., Kaufman, J. (2013) A tale of two tumours: comparison of the immune escape strategies of contagious cancers. Mol. Immunol. 55, 190–193
Solignac, M., Vautrin, D., Baudry, E., Mougel, F., Loiseau, A., et al. (2004) A microsatellite-based linkage map of the honeybee, Apis mellifera L. Genetics 167, 253–262
Solignac, M., Mougel, F., Vautrin, D., Monnerot, M., Cornuet, J.-M. (2007) A third-generation microsatellite-based linkage map of the honey bee, Apis mellifera, and its comparison with the sequence-based physical map. Genome Biol. 8, R66
Sumner, S., Keller, L. (2008) Social evolution: reincar- nation, free-riding and inexplicable modes of repro- duction. Curr. Biol. 18, R206–R207
Suomalainen, E., Saura, A., Lokki, J. (1987) Cytology and evolution in parthenogenesis. CRC Press, Boca Raton, FL
Tiedemann, R., Moll, K., Paulus, K.B., Schlupp, I. (2005) New microsatellite loci confirm hybrid origin, parthenogenetic inheritance, and mitotic gene conversion in the gynogenetic Amazon molly (Poecilia formosa). Mol. Ecol. Notes 5, 586–589
Tribe, G.D. (1983) What is the Cape bee? South African Bee Journal 55, 77–87
Verma, S., Ruttner, F. (1983) Cytological analysis of thelytokous parthenogenesis in the Cape honey bee Apis mellifera capensis. Apidologie 14, 41– 57
Visscher, P.K. (1989) A quantitative study of worker reproduction in honey bee colonies. Behav. Ecol. Sociobiol. 25, 247–254
Weinberg, R. (1998) One renegade cell: the quest for the origins of cancer. Phoenix, London
Wenseleers, T., Alves, D.A., Francoy, T.M., Billen, J., Imperatiz-Fonseca, V.L. (2011) Intraspecific queen parasitism in a highly eusocial bee. Biol. Lett. 7, 173–176
Wheeler, W.M. (1911) The ant-colony as an organism.
Morphology 22, 307–325
White, M. (1984) Chromosomal mechanisms in animal reproduction. Bull. Zoo. 51, 1–23
Winston, M.L. (1991) The biology of the honey bee.
Harvard University Press, Harvard
Wirtz, P., Beetsma, J. (1972) Induction of caste differ- entiation in the honey bee (Apis mellifera) by juvenile hormone. Entomol. Exp. Appl. 15, 517–520
Woyke, J. (1963) What happens to diploid drone larvae in a honeybee colony? J. Apic. Res. 2, 73–75
Zchori-Fein, E., Gottlieb, Y., Kelly, S.E., Brown, J.K., Wilson, J.M., et al. (2001) A newly discovered bacterium associated with parthenogenesis and a change in host selection behavior in parasitoid wasps. Proc. Natl. Acad. Sci. USA 98, 12555– 12560
https://cloverhoney.web.id/
https://cloverhoney.web.id/clover-honey-madu-hdi/
https://cloverhoney.web.id/propoelix/
https://cloverhoney.web.id/royal-jelly-hdi/
https://cloverhoney.web.id/clover-honey-harga/
https://cloverhoney.web.id/propoelix-harga/
https://cloverhoney.web.id/hdi-propoelix-adalah/
https://cloverhoney.web.id/manfaat-propoelix/
https://cloverhoney.web.id/madu-hdi-harga/
https://cloverhoney.web.id/propoelix-plus/
https://cloverhoney.web.id/madu-hdi-manfaatnya/
https://cloverhoney.web.id/clover-honey-manfaatnya/
0 notes
Text
Replying and debate
@luzonmfjel asked:
Hey! About your quirk genetics post, it was a really interesting read on a topic I've been thinking a lot about too. Now all of my genetics qualifications come down to 9th grade biology so this probably doesn't make much sense, but I was wondering if it was possible that quirks were passed on like blood types, with multiple different alleles existing? Not about if you have/don't have a quirk, but more about what that quirk would be. For example, Todoroki. Say his dad’s genotype was Aa, and his mom’s was Bb. With A being fire and B being ice, of course. So put on a punnett square, the possible outcomes would be Ab, Ba, and AB. Ab would mean either a partial or full dominance of fire, Ba of ice. That would result either in someone having a quirk that’s entirely one, or mostly one, like Shouto’s siblings, and AB would be the half and half codominance that we see with him. Those are just the genotypes though, and the way that those genes would present themselves in phenotype could just vary. I'm sorry if that was less than coherent, just thought I'd offer my two cents. ^^
Hell yeah, debate debate. I love so much <3
Don’t fret at all, mate. Let’s see if I understood you right. Like as I see it, you said pretty much the same thing I did (?). It’s just that you chose to use different letters instead of only A’s. Using A’s was just a personal choice, not an avoidance of distinction between quirks. When I described Todoroki’s case... well here, let me just reiterate:
One way it could go is that if a child ends up being dominant homozygous (AA), then they should probably inherit “both” the quirks - and how those genes would express themselves is another point entirely. It could also happen (perhaps more rarely) with the child being heterozygous (Aa), in which one of the parent’s quirk is recessive homozygous (aa). So here are the options:
Mother: Aa
Father: Aa
Possibilities: AA (25%), Aa (50%), aa (25%)
Or:
Mother/father (vice versa): Aa, aa
Possibilities: Aa (50%), aa (50%)
In that description, for Todo’s case, the father “A” would mean Fire, and the mother “A” would mean Ice (or their aa would mean Fire/Ice on the second choice). The possibilities are the punnett square results of that. Besides that I fail to see what the argument here is, as it seems like a pretty good sum up of what I described on that meta. Like, what you said is exactly what I meant in it, so I do apologize if it wasn’t clear and made it sound like I was talking again about quirk vs no-quirk (the intention was to had moved on from the quirkless argument by this point of the meta). I tried not to say which was Todoroki’s genotype, because I wanted to leave it more open, and give more freedom for the whole thing (it had inferences and, in the end, it’s a fiction thing).
However, if you do want to get very realistic I reckon that the correct answer for Todoroki would be that he is an heterozygous (Aa). And the second option is the correct one for the parents. That’s because, as far as I know, Codominance is usually a trait observed on heterozygous. Whereas Partial Dominance is a trait usually observed in homozygous (AA/aa).
About the multiple alleles point (ABO blood type)... That’s not the direction I chose on my Meta, because I thought would make it a bit overcomplicated, and also because I didn’t think it likely that quirks have multiple alleles (which doesn’t mean is not the case). But determining factors for quirk inheritance can involve maybe more than one pair of chromosomes, as well.
I hope this helps, I mean, assuming that’s what you wanted to know. Let me know if you meant something else /o/
@masterlazywriter reblogged your post and added:
Hi! I want to say that I am a biology major and your genetics analyses are on point! I’m very surprise your include epigenetic at the end! :o cause I was thinking about epigenetic for a while while I read it.
THANK YOU! Epigenetic has logic for this situation, doesn’t it? <3 so interesting.
@roquelg replied toy our post “Quirks and Genetics”:
Oh, I love your analysis. It is very well structured. Only one question, aizawa is able to neutralize the alpha factor (the ability to use a quirk) the bullets are the same or do they work also eliminating the quirk? ... Thanks for sharing your ideas
Oh thank you, mate! I’m that glad you liked it! <3 took some time and effort, so this feedback makes me quite happy actually :3
I do need to disclaim that the logic I used for the meta regarding the quirk factor is the hugest leap I did, the largest inference, and it may very well be wrong (regarding the relationship between quirk, quirk factor, plus alpha. I mean, the explanation was confusing and it may be that these 3 terms are interchangeably talking about the same thing, for example. There are other hypothesis as well, but I needed to settle for one to move on with the rest of what I wanted to say).
So, Aizawa says that Erasure “doesn’t attack the quirk itself”, then proceeds to say that Plus Alpha is a mechanism added to a normal human body, and that the collection of these are called Quirk Factor. And then he says that Erasure halts the Quirk Factor. I took that to mean that he affected the response between the Plus Alpha and the Quirk. A friend interpreted it as a neurotransmitter inhibitor and I think that’s a pretty accurate metaphor to how Erasure seems to work. If my hypothesis is correct, that is.
The bullets are a whole other problem. So, in the beginning we are led to believe that they cause damage to the Quirk Factor. In Mirio’s case, a permanent one. However, we now know that Eri’s quirk is that of Rewind, not really the same thing as a quirk that erases quirks. It seems that the bullet formula is using Eri’s blood/quirk to attack specifically the Quirk Factor and Rewind only that part of the body. In theory, the same aspect can be rewinded back to before the person took the bullet. So I wouldn’t say that it’s the same as eliminating the quirk per se.
Applying my theory, it would seem that the bullets rewind the Plus Alpha mechanism to either a time in a person’s life before they developed enough to be able to use their quirks (4 years old in canon), OR to a time in evolution where the Plus Alpha did not exist yet (?). This is all highly hypothetical though, so have that in mind. I hope this answered your question XD let me know if it didn’t.
@sarahhaley101 replied to your post “Quirks and Genetics”:
This analysis is super good and thorough!! However I’m not sure that Todoroki’s quirks is the perfect example of typical codominance. It is not just his quirk that has two expressions — his hair color and eye color do as well, and these multiple alleles at different foci are split right down his center. He could be a chimera! So, he has two completely different sets of chromosomes, not just two alleles at one foci. Chimerism has never been recorded in humans to this degree, but this is anime so!
Thank you so much, mate! <3 This means a lot, I’m very happy with the positive response *was lowkey expecting to receive hate* /o/
Well yeah, but codominance is an expression on different cromossomes and in fact hair colour can have a codominance expression (and todo hair seems to be that). All in all he’s all a codominance isn’t he? I would like to point out that heterochromia is not a result of codominance though. So have that in mind.
All in all it might as well be chimerism, so I can’t say you are wrong. I mean, this is an anime afterall. I just tried to bring real world genetics to the whole thing so talking about that didn’t even cross my mind, to be perfectly honest. It is a thing to consider, of course. Thanks for introducing the idea, mate :3
@bakushima-ima replied to your post “Quirks and Genetics”:
Wait Bakugou was born out of a quirk marriage??? Did I miss this in the manga?????
Awesome analysis though, I love seeing people analyse the scientific side of bnha because it's so well thought out and planned 👍
Why, thank you <3 I’m really glad you enjoyed it, thank you kindly for letting me know! o/
It’s not stated anywhere in canon that Bakugou was born out of a Quirk Marriage. Like, I use the term quirk marriage just to mean the same as quirk fusion. In canon the term “quirk marriage” means a social behaviour in which people marry just so their offspring would have their quirk combined. The fandom has been using that word to also mean a “fused quirk”, so the quirk itself is married, say. It’s just a collective choice, really. So Bakugou’s quirk is a “marriage” between his parent’s quirks, but his parent’s marriage was not a result of a quirk marriage objective (as far as we know). Two different things, then, yeah?
I hope this was your question XD let me know if that was not the case and I misunderstood :3
#bnha meta#boku no hero academia#my hero academia#i hope this is okay it felt easier to just do a big thing instead of answering individually and clogging my followers dashboard#the positive response was so nice#thank you so much#i love yall#amethyst is nerdying out again#bnha chatter#not a request#meta
17 notes
·
View notes
Text
Behavioural medicine journals- BJSTR Journal

Diagnostic Work Up in A Patient with A Progressive Cardio-/Encephalomyopathy, Peripheral Neuropathy and Respiratory Stridor by Manuela Zlamy* in Biomedical Journal of Scientific & Technical Research (BJSTR) https://biomedres.us/fulltexts/BJSTR.MS.ID.001209.php#
Introduction: Mitochondrial disorders have a clinical and genetic heterogeneity which often complicates the diagnostic process especially in critically ill patients. Exome sequencing can help to generate a definite diagnosis in a rather short time.Case Presentation: A boy presented with inspiratory stridor, respiratory insufficiency, progressive muscular hypotonia, poor head control and sucking, and hypersomnia at three months of age. He got ventilator dependent and developed cardiomyopathy within weeks. The progressive multisystem disease leads to the suspicion of a mitochondrial disorder. As neither the clinical nor the metabolic changes or the diagnostic procedures performed seemed indicative of a specific mitochondrial disorder, we performed exome sequencing. This revealed a known homozygous mutation (c.418G>A, p.Glu140Lys) in SCO2.Discussion: Retrospectively, the combination of progressive encephalomyopathy and inspiratory stridor could have been the diagnostic clue for SCO2 deficiency. However, given the enormous clinical and genetic heterogeneity it is nearly impossible to diagnose a mitochondrial disorder with single gene analysis in the restricted time frame clinicians’ face when caring for life- threatening ill infants.
For more Articles on Behavioural medicine journals please click here https://biomedres.us/index.php
For bjstr journal
#biomedical open access journals#bjstr journal#Journals on Psychology#Nuereological disorders#Immunological diseases
0 notes
Text
Command Parenting Manual.
The loved ones's story was published to the WTOC Facebook page on Monday and also has actually already gotten over 11,000 likes as well as lots of reviews, usually on behalf of Mr. Schlichenmeyer. The parent shares to the youngster that they take exactly what the pleased end of the tale means when moms and dads share fairy stories with their children. So if it's on the spine of my phone, I am actually going to get a visual hint to advise me and keep that leading of mind so I keep paid attention to that.
Select the PSST company logo in the best advertisement of this particular page, to make certain you get on the PSST home-page, before using this search option. Building your private brand name generates genuineness, attraction, possibilities, as well as improved power (due to the fact that aiming to be exactly what you think a particular viewers wishes has a great deal of electricity).
Even if a parent struggles with Alzheimer's or even mental deterioration, think about your own self as an overview of assisting them in their ultimate years. More mature youngsters might even enter the abuse to guide the attention into on their own and off of the struggling parent.
Probably these will certainly inspire all kind of cheery imagination for a terrific party from the little X-mas elf in you! If you're a moms and dad which carries out some of these things, I hope you'll stop that behaviour straightaway. Occasionally, the remittance of little one help is actually additionally proportional to the time the kid devotes with each parent.
In most American conditions, courthouses choose approving joint legal protection of the child, unless the other moms and dad declines custody, or is actually otherwise regarded as unhealthy. However, lots of moms and dads and also kids are uninformed this this wonderful, enjoyable, leading 10 X-mas present as you could only discover this at particular shops.
Yes, there is a long road but possessing a private self-supporting grownup can be obtained to the advantage to each moms and dad and also grown-up youngster. Complying with a separation lots of find yourself delivering youngsters up as a solitary parent which can be demanding and tough.
If a homozygous dominant moms and dad possesses a child along with a homozygous dormant parent at that point all the little ones will definitely made the left thumb over the right. The online search engine and also analysis devices illustrated here are ones I have actually utilized as an instructor and parent with my children, in the class and in your home.
Teach regarding a number of the terrific entertainers Top-diet-products.info Elvis Presley, Bob Dylan, Madonna, as well as Michael Jackson, that were actually leading favorite performers of the past with to the here and now. I was actually a singular parent therefore selection making regarding all problems fell to me. You are actually in the very same scenario if you are a single moms and dad.
It felt really good even when this is actually not a totally equivalent reveal and most importantly this was what I experienced my parent will wish me to carry out to fulfill their final wishes. You may receive definitely imaginative with that said idea as well as go wild along with creating this looks great.
Actually, the majority of little ones stemming from singular parent families grow with a huge amount of appreciation for their moms and dad. Some classrooms also include an anonymous moms and dad comment box away from their classroom. China's NIKE National Junior Ping Pong Rank Competition belongs from Nike Junior Scenic Tour.
0 notes
Text
Putting attention in the spotlight: The influence of APOE genotype on visual search in mid adulthood
Publication date: 15 September 2017
Source:Behavioural Brain Research, Volume 334
Author(s): Claire Lancaster, Sophie Forster, Naji Tabet, Jennifer Rusted
The Apolipoprotein E e4 allele is associated with greater cognitive decline with age, yet effects of this gene are also observed earlier in the lifespan. This research explores genotype differences (e2, e3, e4) in the allocation of visuospatial attention in mid-adulthood. Sixty-six volunteers, aged 45–55 years, completed two paradigms probing the active selection of information at the focus of attention (a dynamic scaling task) and perceptual capacity differences. Two methods of statistical comparison (parametric statistics, Bayesian inference) found no significant difference between e4 carriers and the homozygous e3 group on either the dynamic scaling or perceptual load task. E2 carriers, however, demonstrated less efficient visual search performance on the dynamic scaling task. The lack of an e4 difference in visuospatial attention, despite previous suggestion in the literature of genotype effects, indicates that select attentional processes are intact in e4 carriers in mid-adulthood. The association of e2 genotype with slower visual search performance complicates the premised protective effects of this allele in cognitive ageing.
from #ORL-Sfakianakis via xlomafota13 on Inoreader http://ift.tt/2uinaF8
from OtoRhinoLaryngology - Alexandros G. Sfakianakis via Alexandros G.Sfakianakis on Inoreader http://ift.tt/2tYD0oU
0 notes
Text
AAPA Presentation, by Sam Larson: “An evolutionary perspective on the contribution of serotonergic genetics to health: lessons from rhesus macaques”
SAM M. LARSON1, ANGELINA RUIZ-LAMBIDES2, JULIE HORVATH3, ATHY ROBINSON4, PATE SKENE4, MICHAEL L. PLATT5,6,7 and LAUREN J.N. BRENT8
1Department of Anthropology, University of Pennsylvania, 2Caribbean Primate Research Center, University of Puerto Rico, 3Biological & Biomedical Sciences, North Carolina Central University, 4Department of Neurobiology, Duke University, 5Department of Neuroscience, Perelman School of Medicine, University of Pennsylvania, 6Marketing Department, The Wharton School, University of Pennsylvania, 7Department of Psychology, University of Pennsylvania, 8Centre for Research in Animal Behaviour, University of Exeter
Genomic variation related to serotonin biosynthesis can serve as risk factor for psychopathology, particularly when following early-life stress. Recent evolutionary informed theory posits that this relationship, rather than uniformly maladaptive, may be advantageous given certain social contexts. Here, we examine mortality trajectories according to serotonergic genotype in a free-ranging nonhuman primate (Macaca mulatta). We fit life-tables to subjects genotyped for VNTR length polymorphisms—short (S) vs. long (L)—of two serotonergic genes (5-HTT and TPH2). Females carrying at least one copy of the less frequent 5-HTT(S) and TPH2(L) alleles had greater mortality rates in early life but slower rates of senescence, and thus ultimately lived longer than females homozygous for 5-HTT(L) or TPH2(S)respectively. For 5-HTT, an opposite pattern is obse rved in males, where those carrying at least one copy of the 5-HTT(S) have shorter lifespans than homozygous 5-HTT(L) males. Males TPH2(s)homozygotes, like the females, have a reduced life expectancy than those carrying a copy of TPH2(L). Our results are consistent with the hypothesis that poor functioning of the serotonergic system results in increased sensitivity to the social environment, which may better serve female macaques than male. We discuss our findings in light of widely reported sex differences in sensitivity to stress, development of psychopathologies, and resilience in the context of social support, especially as mediated by serotonergic genetics. As mortality is a major component of fitness, this study offers some resolution to the question of the persistence of serotonergic allelic variation in human evolution.
#anthropology#primatology#genetics#adaptation#stress#social behavior#mortality#fitness#conference#aapa#announcements
0 notes
Text
Altered Striatal Synaptic Function and Abnormal Behaviour in Shank3 Exon4-9 Deletion Mouse Model of Autism.
PubMed: Altered Striatal Synaptic Function and Abnormal Behaviour in Shank3 Exon4-9 Deletion Mouse Model of Autism.: Related Articles Altered Striatal Synaptic Function and Abnormal Behaviour in Shank3 Exon4-9 Deletion Mouse Model of Autism. Autism Res. 2016 Mar;9(3):350-75 Authors: Jaramillo TC, Speed HE, Xuan Z, Reimers JM, Liu S, Powell CM Abstract Shank3 is a multi-domain, synaptic scaffolding protein that organizes proteins in the postsynaptic density of excitatory synapses. Clinical studies suggest that ∼ 0.5% of autism spectrum disorder (ASD) cases may involve SHANK3 mutation/deletion. Patients with SHANK3 mutations exhibit deficits in cognition along with delayed/impaired speech/language and repetitive and obsessive/compulsive-like (OCD-like) behaviors. To examine how mutation/deletion of SHANK3 might alter brain function leading to ASD, we have independently created mice with deletion of Shank3 exons 4-9, a region implicated in ASD patients. We find that homozygous deletion of exons 4-9 (Shank3(e4-9) KO) results in loss of the two highest molecular weight isoforms of Shank3 and a significant reduction in other isoforms. Behaviorally, both Shank3(e4-9) heterozygous (HET) and Shank3(e4-9) KO mice display increased repetitive grooming, deficits in novel and spatial object recognition learning and memory, and abnormal ultrasonic vocalizations. Shank3(e4-9) KO mice also display abnormal social interaction when paired with one another. Analysis of synaptosome fractions from striata of Shank3(e4-9) KO mice reveals decreased Homer1b/c, GluA2, and GluA3 expression. Both Shank3(e4-9) HET and KO demonstrated a significant reduction in NMDA/AMPA ratio at excitatory synapses onto striatal medium spiny neurons. Furthermore, Shank3(e4-9) KO mice displayed reduced hippocampal LTP despite normal baseline synaptic transmission. Collectively these behavioral, biochemical and physiological changes suggest Shank3 isoforms have region-specific roles in regulation of AMPAR subunit localization and NMDAR function in the Shank3(e4-9) mutant mouse model of autism. PMID: 26559786 [PubMed - indexed for MEDLINE] http://dlvr.it/N5fn78
0 notes
Last Seen Blogs

miky-rei
Two Miku kissing each other

cazass
Poking my brain with a stick till it moves

sissybridewifeandmother
sissy housewife

ilovezoobookss
giant yaoi butt

circustanflautico
circustanflautic-o