#the fossil was actually a dinosaur. But I tried to keep these fairly well-known.
Note
Hide 2, kimi 2, 7, and 13 for any au!
Alright! Pardon the wait,
2. Hide headcanon,
Collects those little eraser animal things and hands them out like candy. You're having a bad day? Eraser zebra on your desk. You're proud of yourself? Eraser penguin appears in your jacket pocket. Confused and stressed? Eraser lion. Do not ask when he put them there, he'll just dodge the question and use it as a transition to talk about why he gave them to you. The secrets of his stealth are unfathomable to any mere mortal, just accept your new friend and let it bring you strength.
2. Kimi headcanon!
Impressively talented at arcade games, she will kick your ass. It's a combination of knowing all the tricks to beating their scams and raw skill. Nishiki had a very rude awakening to this when he had this whole grand plan for a nice date, where he was going to win her something from the claw machine and then they'd get icecream together. Only to witness these plans crumble as he was beaten 6 times in pinball, and witnessed her break the laws of physics to grab exactly the stuffed animal she wanted from the claw machine 3 times in a row without dropping them.
She gave him one and he's a little bit afraid of it.
7. Angst headcanon
I'm not a very angst-ish person by nature regardless of what my fics/drawings might suggest but I'll try my best...
Rotten Follow is a quinque made from Shikorae's deceased older brother, something that undeniably has extremely polarizing emotions tied to it because on one hand the memory of his brother seems to be the only thing he's managed to hold on to, but he was also tortured into a catatonic state with that quinque and only kept alive by the small bits of his brother's body he was fed over the years which is YIKES even for Tokyo Ghoul. As much as I love Shikorae I do think he is a prime example of the prioritization of "dark" subjects over logic as well as several other issues with the portrayal of villainous ghouls in the story.
Anyways, with all this context in mind I don't think it's too much of a reach to say that Shikorae's rampage could've been motivated by the inheritor of Rotten Follow after Furuta (Whoever that could be) and a simple desire to chase after any trace of his brother. Because he's never been shown to be explicitly malicious (given he hasn't been shown as the contrary either) simply following orders and hunger.
13. A headcanon about a AU (of your preference)
Not great at AU's either because I'm mostly fond of minor alterations that have ripple effects and no one wants to hear me ramble for 3 paragraphs about how if Furuta lived who's to say his RC system couldn't get worse actually- anyways may's just around the corner so mermaid AU, exclusively with some of my favorites and also Arima:
Hanbee would be a False Killer Whale
Juuzou would be a Bottlenose (They form partnerships sometimes it's really interesting)
Hairu's a River Dolphin.
Arima's a Sperm Whale.
Rize's a Giant Pacific Octopus or Great White.
Furuta's a Mimic Octopus.
Koori's a Blue Pinoy Angelfish.
Uta's a Killer Whale
Renji's an Angel Shark or Thresher
Itori's a mantis shrimp
#I know an obnoxious amount about sea life actually because I went through a really aggressive phase as a child#where I wanted to be a an underwater archaeologist and I was convinced I needed to know all the current sea life so I'd be certain#the fossil was actually a dinosaur. But I tried to keep these fairly well-known.#miscellaneous not-art things#zeph answers questions about media/characters/ships#tokyo ghoul
8 notes
·
View notes
Text
Deinonychus antirrhopus

By Ripley Cook
Etymology: Terrible Claw
First Described By: Ostrom, 1969
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Theropoda, Neotheropoda, Averostra, Tetanurae, Orionides, Avetheropoda, Coelurosauria, Tyrannoraptora, Maniraptoromorpha, Maniraptoriformes, Maniraptora, Pennaraptora, Paraves, Eumaniraptora, Dromaeosauridae, Eudromaeosauria, Dromaeosaurinae
Status: Extinct
Time and Place: From 115 to 98 million years ago, from the Aptian of the Early Cretaceous until the Cenomanian of the Late Cretaceous
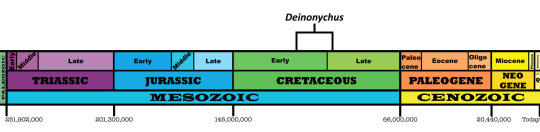

Deinonychus is known from the Little Sheep and Himes Members of the Cloverly Formation of Montana and Wyoming, the Middle Member of the Antlers Formation of Oklahoma, and the Ruby Ranch and Mussentuchit Members of the Cedar Mountain Formation of Utah.

Physical Description: Deinonychus is a Dromaeosaur, or “raptor” dinosaur, and one of the best known of this group. Like other raptors, it looked a lot like a modern bird - fully covered in feathers, with large wings, a fan of feathers on the tail, and a triangular head. However, unlike a living bird, it was unable to fly, did not have a beak, and had a bony tail. Raptors are also characterized by their huge sickle claws on their feet, positioned on the second toe.
By Dinoguy2, CC BY-SA 2.5
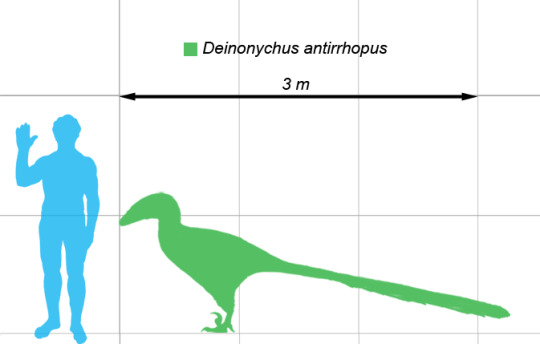
Deinonychus specifically was fairly middle of the road in terms of raptor size, reaching up to 3.4 meters in length, and only coming up to about a meter in height. It would have been fairly lightweight, due to the air sacs present in the bones (like living birds). This would have made it about the size of a small wolf or a large dog - still smaller than the raptors in Jurassic Park. It had large openings in the skull to help keep it lightweight, and these were larger than those of other raptors; and it also had a more robust skull roof compared to other raptors. It had a long, narrow snout, which flared rapidly at the eye portion of the skull to give Deinonychus 3D vision.
Deinonychus had a long body, with a relatively short torso and extremely long tail. This tail was stiffened, to keep it very straight and only allowing a little bit of side to side movement. In addition, Deinonychus had a pelvis like a modern bird, giving more room to the gut for digesting food. As for limbs, Deinonychus had particularly long bones in its wing, which would have supported large wing feathers, much like wings of birds today. This is supported by the fact that most raptors found with feather impressions - Microraptor and Zhenyuanlong, notably - have full sized wings. Deinonychus also had a large fan of feathers across the tail. The legs of Deinonychus were fairly short compared to the rest of its body. It would have been warm-blooded, and an extremely active, birdy animal.

By Emily Willoughby, CC BY-SA 4.0
Diet: Deinonychus was a meat-eater, and a predator to boot, associated with animals such as Tenontosaurus and other medium-sized herbivores, though it probably also ate smaller animals such as mammals as well.
Behavior: Deinonychus has been the subject of extensive debate on the behavior of predatory dinosaurs, due to the fact that a large number of Deinonychus were found together with the herbivorous Tenontosaurus. This lead to the natural conclusion that Deinonychus was feeding on Tenontosaurus, and that the group had brought down the prey together - as a pack.

By José Carlos Cortés
This has sparked a lot of controversy, as Deinonychus are significantly smaller than Tenontosaurus, so one taking down this dinosaur alone is rather unlikely. However, most archosaurs don’t actually do cooperative hunting - though crocodilians and living birds might work together on bringing down food, it isn’t usually done in any coordinated fashion, but rather out of convenience. This lead to the idea of pack hunting not being particularly likely in Deinonychus, since living dinosaurs don’t really do much of it either. However, there are birds of prey, as well as crocodilians, that show gregarious pack hunting behavior, though not anything to the complexity seen in mammals. In addition, there are trackways that are parallel, with similar pacing of footprints, indicating that Deinonychus were living in social groups that traveled together. This would have allowed them to bring down larger prey.
In addition to this, Deinonychus probably utilized raptor prey restraint to hold down prey. Raptor Prey Restraint, or RPR, is a behavior seen today in living birds of prey, that would have been possible for Deinonychus and other Dromaeosaurs to do. With this method, Deinonychus would pin down it’s prey with its foot claws and, while holding it down, flap its wings rapidly in order to keep balanced as the prey moved around and tried to escape beneath it. This would have allowed Deinonychus to feed on the prey while it was still alive. This is supported not only by Deinonychus having long arms to support large wings, but also by its foot claws being similar to those of living owls, giving strong foot strength for holding onto prey.

By Tas on i-draws-dinosaurs
It’s entirely possible, therefore, that Deinonychus could have grouped up with others of its kind - probably a family grouping - and attacked herbivores much larger than itself, with some members of the group leaping onto the backs of these herbivores and holding on using RPR. The large sickle claws were not quite built for slashing at the prey, but rather for pinning down the prey and stabbing strategically at the animal, allowing it to bleed out rapidly. In addition, Deinonychus could have used the claws on its hands for slashing when it wasn’t using the wings for flight, as well as for grasping food. Like the modern seriema, however, Deinonychus could probably have torn apart smaller prey animals with its feet when eating a solo meal, if necessary.
Though raptors are commonly portrayed as being extremely fast (to quote a movie, “cheetah speed”), this is unlikely given the length of the bones of the leg in Deinonychus and other raptors. It had short bones in its foot as well as its leg, indicating that it was not very fast at running, and probably would have been more of an ambush than a pursuit predator. It might have been very good at leaping, however, and could have used its wings to generate a little lift to take the jump farther, and pounce on its food. Thus, Deinonychus could leap onto large prey such as Tenontosaurus, to get into a position to use RPR and feed upon it. The strong bones on the foot would have aided in handling the stress from such a jump. Deinonychus would have also been able to use its jaws in taking down prey, able to bite through bone if needed with a very high bite force. However, it did not do this habitually, like Tyrannosaurus, and instead used this bite force for defense and the capture of prey, rather than feeding.

By Ashley Patch
Though Deinonychus was too large to fly, it could generate enough lift to utilize another behavior of living birds - wing-assisted incline running, or WAIR. WAIR involves the flapping of wings fast enough that, with a hop and enough speed in the hind limbs, a bird is able to run up a steep surface such as a tree trunk, a wall, or a cliff face. Deinonychus might have been able to use WAIR to run up the surfaces of trees or after prey going up a tall feature such as a mountain and a hill - while Deinonychus was not a fast runner, WAIR could have helped it to gain some lift in these situations and go faster up the mountain or hill to catch its prey. Deinonychus also had a very stiffened tail, which would have helped it to keep its balance as it utilized WAIR and RPR in hunting and movement.
All in all, these traits - family groups, ambush predators, bursts of speed rather than long periods of speed, and pouncing - indicate that Deinonychus and other raptors like it were more like bird-cats, rather than bird-dogs, in terms of similar ecologies and behavior. You know what would have been bird-dogs? Tyrannosaurs. But more on that another time.
Interestingly enough, juvenile - young - Deinonychus are a bit different in proportion than the adults, namely by having longer arm bones and more sharply curved claws. This might mean that juvenile and baby Deinonychus actually went into trees, for safety or to help get food. It is even possible that young Deinonychus could fly, and grew out of it as they got bigger, but that is a hypothesis that has generated quite a bit of controversy and requires more extensive research.

By Kevin on Jurasicore
Deinonychus took care of its young, and eggs have been associated with Deinonychus fossils. Deinonychus would have brooded its eggs like living birds, protecting the nest with its large wings and sitting on top of the nest to aid in keeping the eggs warm and protected. It’s possible that the family groups of Deinonychus would have worked together to protect the nest until the young hatched. We know that Deinonychus laid blue eggs, helping to camouflage them in the foliage and against predators, a trait that evolved in Deinonychus and stuck around through modern birds. Deinonychus young then probably would escape to trees and other high locations whenever the parents weren’t around to protect them; as they got bigger they were probably able to get their own food, but they could have worked together to get large herbivores such as Tenontosaurus.
As we have seen, Deinonychus had a lot of reasons to have the large wings - for hunting, keeping itself and its young warm, and for movement. In addition to this, however, it’s likely that the wings were used for display - and, indeed, this is probably why complex feathers evolved in the first place. These large wings would have been perfect for complicated patterns and colors, which could have then been shown off to other raptors in mating displays. The large tail fan would also have been a good display structure, and could have been lifted and raised in situations of communication between members of the species, as well as in mating displays. By fluffing out its wings and raising up its tail, Deinonychus would have made itself look much bigger, which could have been used to express that it was threatening another animal, or that it was looking for sex. This is, of course, speculative, but based on behavior of living animals with similar structures.
In short, this was a medium-sized, active and agile predator, extremely birdlike in both appearance and behavior. It probably showed fairly complicated social behavior, and utilized its wings for complicated predatory actions. This makes Deinonychus - and other raptors - important keystones in our understanding of the complexity of Mesozoic dinosaurs and the evolution of birds.

By Scott Reid
Ecosystem: Deinonychus is known from the distinctive environments of the “middle” Cretaceous of Western North America, a snapshot between the iconic ecosystems of the late Jurassic and the late Cretaceous - indeed, many of the same environments as Sauropelta, who we learned about earlier this week.
In the Cloverly Formation, Deinonychus lived alongside the Western Interior Seaway as it grew through North America, and the dinosaurs present in this ecosystem such as Deinonychus would have been associated primarily with the floodplain and river system there. The flooding rivers caused mass burials of animals, and then eventually fossil formation. The plains were forested and covered with a variety of conifers and cycads. Deinonychus is known from both the Little Sheep and Himes Members of the Cloverly Formation, and thus was able to persist in this environment even as the Western Interior Seaway grew, and it turned from muddy to more sand based.
In the earlier, muddier Little Sheep ecosystem, Deinonychus lived alongside the ankylosaur Sauropelta, the ornithopod Tenontosaurus, the fast-moving Zephyrosaurus, the oviraptor Microvenator, and an unnamed large sauropod. There were also lungfish, sharks, and many bony fish, frogs, salamanders, and turtles. Mammals were also extremely common in this ecosystem and were probably a good source of prey for Deinonychus - animals such as Atokatheridium, Gobiconodon, Oklatheridium, and Paracimexomys. There were also plenty of crocodilians and lizards.

By Fred Wierum, CC BY-SA 4.0
In the later, sandier Himes ecosystem, Deinonychus still lived with Microvenator, Sauropelta, and Tenontosaurus, but also the large sauropod Sauroposeidon and the poorly known sauropod Rugocaudia, the nodosaur Tatankacephalus, and the larger predator Acrocanthosaurus which would have been a major threat to Deinonychus. There was also potentially a proto-bird in the ecosystem. There were many different kinds of lizards, frogs, salamanders, crocodylomorphs, turtles, fish, and sharks, as well as many mammals for Deinonychus to eat in addition to Tenontosaurus.
Deinonychus is also known from the Antlers Formation, from slightly more east in North America. It is known from the middle member of this ecosystem, and from about the same time as the Cloverly Formation to the west. This was also a floodplain ecosystem, which was periodically flooded with sources of water. Here, there was the tall sauropod Sauroposeidon, the ornithopod Tenontosaurus, a variety of mammals such as Astroconodon and Atokatheridium and Oklatheridium and Pappotherium, sharks and bony fish, and many types of Crocodylomorphs. There was also the large predator Acrocanthosaurus which, again, probably would have been a major predator of Deinonychus. Unfortunately, no plant fossils are known from the Antlers Formation to better piece together details as to the foliage Deinonychus would have lived among.
Finally, Deinonychus is also known from the Cedar Mountain Formation, which was an ecosystem that immediately followed the famous Morrison Formation of the Late Jurassic. Deinonychus is known from later time periods in the Cedar Mountain Formation than right after the Morrison, of course, but this ecosystem does represent a transitory time period between the Late Jurassic and Late Cretaceous. Here, Deinonychus is known from the Ruby Ranch and Mussentuchit members, which were the two latest occuring ecosystems in the formation. During the Ruby Ranch period - the earlier one - there was an extensive amount of mud, created by a large number of river channels forming due to the growing Western Interior Seaway. Here there were a lot of ferns, magnolias, conifers, and cycads, indicating a mainly forested ecosystem surrounding these rivers. Deinonychus was accompanied by the ankylosaur Gastonia, the ornithopod Tenontosaurus, the sauropods Astrodon and Brontomerus, the large predator Acrocanthosaurus, and a variety of crocodilians.

By Emily Willoughby, CC BY-SA 3.0
The later Mussentuchit Member was still covered in ferns, magnolias, conifers, and cycads, so the actual foliage didn’t seem to change much. However, there does appear to have been a volcanic explosion, leading to more detailed preservation of the animals and plants present - and most fossils from Cedar Mountain come, as such, from this ecosystem. Here, Deinonychus was accompanied by the ankylosaurs Animantarx, Cedarpelta, and Peloroplites - so, unfortunately, Gastonia went extinct. There were other ornithopods besides Tenontosaurus, though Tenontosaurus was present - such as Eolambia and the fast moving Zephyrosaurus. The only known sauropod from the Mussentuchit is Abydosaurus, indicating a drop in sauropod diversity that would continue in North America throughout the Cretaceous. The larger predator Siats was present, as well as the mysterious theropod Richardoestesia and a potential aquatic birdie dinosaur. There was a variety of sharks, bony fish, and lungfish; salamanders and frogs; turtles, snakes, lizards, and crocodilians; and many kinds of mammals present in this ecosystem as well.
Other: Deinonychus was of vital importance to the understanding of the evolution of birds. As a definitively active and bird-like dinosaur, Deinonychus showed researchers in the middle of the twentieth century that dinosaurs were actually the most likely candidates for the ancestors to modern birds. As research went on and more birdie dinosaurs were discovered, this became more and more certain - a path started by Deinonychus. In addition, Deinonychus in conjunction with Maiasaura helped to transition our understanding of dinosaurs from “slow, stupid, big lizards” to “active almost-birds”, a transition in perception that was cemented by the Jurassic Park Franchise. Deinonychus was a fairly derived raptor dinosaur, closely related to some of the largest members of the group such as Achillobator and Utahraptor.
~ By Meig Dickson
Sources under the cut
Adams, D. 1987. The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs. Currie, P. J., E. Koster (ed.). Fourth Symposium on Mesozoic Terrestrial Ecosystems. Tyrrell Museum, Drumheller, Canada: 1 - 6.
Aubrey, W.M. 1998. A newly discovered, widespread fluvial facies and unconformity marking the Upper Jurassic/Lower Cretaceous boundary, Colorado Plateau. Modern Geology, v. 22, p. 209-233.
Barsbold, R. 1983. Carnivorous Dinosaurs from the Cretaceous of Mongolia. The Joint Soviet-Mongolian Palaeontological Expedition Transactions 19: 1 - 117.
Benton, M. J. 2004. Vertebrate Palaeontology (3rd ed.). Blackwell Publishing: 472.
Brinkman, D. L., R. L. Cifelli, N. J. Czaplewski. 1998. First occurrence of Deinonychus antirrhopus (Dinosauria: Theorpoda) from the Antlers Formation (Lower Cretaceous: Aptian - Albian of Oklahoma. Oklahoma Geological Survey Bulletin 146: 1 - 27.
Brinkman, D.B., Scheetz, R.D., Jensen, C., Britt, B.B., and Ortiz, N., 2015, A basal baenid turtle provides insights into the aquatic fauna of the Early Cretaceous (Aptian) Cedar Mountain Formation of west-central Utah. Journal of Vertebrate Paleontology, Abstracts and Program, p. 96.
Brownstein, C. D. 2018. A large Dromaeosaurid from North Carolina. Cretaceous Research 92: 1 - 7.
Burton, D., B. W. Greenhalgh, B. B. Britt, B. J. Kowallis, W. S. Elliott, R. Barrick. 2006. New radiometric ages from the Cedar Mountain Formation, Utah and the Clovelry Formation, Wyoming: implications for contained dinosaur faunas. Geological Society of America Abstracts with Programs 38 (7): 52.
Campione, N. E., D. C. Evans, C. M. Brown, M. T. Carrano. 2014. Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions. Methods in Ecology and Evolution 5 (9): 913 - 923.
Carpenter, K. 1998. Evidence of predatory behavior by carnivorous dinosaurs. Gaia 15: 135 - 144.
Carpenter, K. 2002. Forelimb biomechanics of nonavian theropod dinosaurs in predation. Senckenbergiana Lethaea 82: 59 - 76.
Carpenter, K., 2006, Assessing dinosaur faunal turnover in the Cedar Mountain Formation (Lower Cretaceous) of eastern Utah, USA. Ninth International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstract and Proceedings Volume, p. 21-25.
Carpenter, Kenneth; Bartlett, Jeff; Bird, John; Barrick, Reese 2008. Ankylosaurs from the Price River Quarries, Cedar Mountain Formation (Lower Cretaceous), east-central Utah. Journal of Vertebrate Paleontology. 28 (4): 1089–1101.
Chen, Z.-Q., S. Lubin. 1997. A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming. The Mountain Geologist 34: 51 - 62.
Chure, Daniel; Britt, Brooks; Whitlock, John A.; Wilson, Jeffrey A. 2010. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Naturwissenschaften. 97 (4): 379–91.
Cifelli, R. L. 1997. First notice on Mesozoic mammals from Oklahoma. Oklahoma Geology Notes 57(1):4-17
Cifelli, R. L., J. D. Gardner, R. L. Nydam, D. L. Brinkman. 1997. Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes 57(4):124-131
Cifelli, R.L., Wible, J.R., and Jenkins, F.A. 1998. Triconodont mammals from the Cloverly Formation (Lower Cretaceous), Montana and Wyoming. Journal of Vertebrate Paleontology 18: 237-241.
Cifelli, R.L. 1999. Tribosphenic mammal from the North American Early Cretaceous. Nature 401:363-366.
Cifelli, R. Gardner, J.D., Nydam, R.L., and Brinkman, D.L. 1999. Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes 57:124-131.
Cifelli, R. L., Nydam, R. L., Gardner, J. D., Weil, A., Eaton, J. G., Kirkland, J. I., and Madsen, S., 1999, Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County, Utah: the Mussentuchit Local Fauna, in, Gillette, D., Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication 99-1, p. 219-242.
Currie, P. J. 1995. New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal of Vertebrate Paleontology 15 (3): 576 - 591.
D'Emic, M.D., B.Z. Foreman. (2012). The beginning of the sauropod dinosaur hiatus in North America: insights from the Lower Cretaceous Cloverly Formation of Wyoming. Journal of Vertebrate Paleontology 32(4):883–902.
D'Emic, Michael D.; Melstrom, Keegan M.; Eddy, Drew R. (2012). "Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13–23.
Darton, N. H. 1904. Comparison of the stratigraphy of the Black Hills, Bighorn Mountains, and Rocky Mountain Front Range. Geological Society of America Bulletin 15: 379 - 448.
Davies, S. J. J. F. 2002. Ratites and Tinamous. New York: Oxford University Press.
Delcourt, R., O. N. Grillo. 2014. On maniraptoran material (Dinosauria: Theropoda) from Vale do Rio do Peixe Formation, Bauru Group, Brazil. Revista Brasileira de Paleontologia 17(3):307-316
DePalma, R. A., D. A. Burnham, L. D. Martin, P. L. Larson, R. T. Bakker. 2015. The first Giant Raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. Paleontological Contributions 14: 1-16.
Dinets, V. 2014. Apparent coordination and collaboration in cooperatively hunting crocodilians. Ethology Ecology & Evolution 27 (2): 244 - 250.
Erickson, G. M., K. Curry Rogers, D. J. Varricchio, M. A. Norell, X. Xu. 2007. Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition. Biology Letters 3 (5): 558 - 61.
Evans, D. C., D. W. Larson, P. J. Currie. 2013. A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America. Naturwissenschaften 100 (11): 1041 - 1049.
Fastovsky, D. E., D. B. Weishampel. 2005. Theropoda I: Nature Red in Tooth and Claw. In Fastovsky, D. E., D. B. Weishampel. The Evolution and Extinction of the Dinosaurs (2nd ed.). Cambridge: Cambridge University Press: 265 - 299.
Forster, C. A. 1984. The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana. The MOsasaur 2: 151 - 163.
Fowler, D. W., E. A. Freedman, J. B. Scannella, R. E. Kambic. 2011. The Predatory Ecology of Deinonychus and the Origin of FLapping in Birds. PLoS ONE 6 (12): e28964.
Frederickson, J. A., M. H. Engel, R. L. Cifelli. 2018. Niche Partitioning in Theropod Dinosaurs: Diet and Habitat Preference in Predators from the Uppermost Cedar Mountain Formation (Utah, U.S.A.). Scientific Reports 8: 17872.
Gauthier, J., K. Padian. 1985. Phylogenetic, Functional, and Aerodynamic Analyses of the Origin of Birds and their Flight. Hecht, M. K., J. H. Ostrom, G. Viohl, P. Wellnhofer (ed.). The Beginnings of Birds. Proceedings of the International Archaeopteryx Conference, Eichstätt: Freunde des Jura-Museums Eichstätt: 185 - 197.
Gianechini, F. A., P. J. Makovicky, S. Apesteguía. 2011. The teeth of the unenlagiine theropod Buitreraptor from the Cretaceous of Patagonia, Argentina, and the unusual dentition of the Gondwanan dromaeosaurids. Acta Palaeontologica Polonica 56(2):279-290
Gignac, P. M., P. J. Makovicky, G. M. Erickson, R. P. Walsh. 2010. A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations. Journal of Vertebrate Paleontology 30(4):1169-1177
Gilliard, E. T. 1958. Living Birds of the world. Garden City, NY: Doubleday.
Gishlick, A. D. 2001. The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight. In Gauthier, J., L. F. Gall. New Perspectives on the Origin and Early Evolution of Birds. New Haven: Yale Peabody Museum: 301 - 318.
Grellet-Tinner, G. 2006. Oology and the Evolution of Thermophysiology in Saurischian Dinosaurs: Homeotherm and Endotherm Deinonychosaurians? Papeis Avulsos de Zoologia 46 (1): 1 - 10.
Grellet-Tinner, G., P. Makovicky. 2006. A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications. Canadian Journal of Earth Sciences 43 (6): 705 - 719.
Hill, R.T. 1894. Geology of parts of Texas, Indian Territory and Arkansas adjacent to Red River. Bulletin of the Geological Society of America. 5: 303.
Horner, John R. Dinosaurs Under the Big Sky (Cloverly Formation). Mountain Press Publishing Company. pp. 93–100.
Hwang, S. H., M. A. Norell, Q. Ji, K. Gao. 2002. New Specimes of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China. American Museum Novitates 3381: 1 - 44.
Jenkins, F. A., C. R. Schaff. 1988. The Early Cretaceous Mammal Gobiconodon (Mammalia, Triconodonta) from the Cloverly Formation in Montana. Journal of Vertebrate Paleontology 8(1):1-24
Kielan-Jarorowska, Z., and Cifelli, R.L. 2001. Primitive boreosphenidan mammal (?Deltatheroida) from the Early Cretaceous of Oklahoma. Acta Palaeontologica Polonica 46: 377-391.
Kinneer, B., K. Carpenter, A. Shaw. 2016. Redescription of Gastonia burgei (Dinosauria: Ankylosauria, Polacanthidae), and description of a new species. Neues Jahrbuch Geologie und Paläontologie, Abhandlungen. 282 (1): 37–80.
Kirkland, J.I., Britt, B., Burge, D., Carpenter, K., Cifelli, R., DeCourten, F., Eaton, J., Hasiotis, S., and Lawton, T., 1997b, Lower to Middle Cretaceous dinosaur faunas of the Central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution, and biogeography. Brigham Young University Geology Studies, v. 42, p. 69-103.
Kirkland, J.I. and Madsen, S.K. 2007. The Lower Cretaceous Cedar Mountain Formation, eastern Utah: the view up an always interesting learning curve. Fieldtrip Guidebook, Geological Society of America, Rocky Mountain Section. 1-108 p.
Kofron, C. P. 2003. Case histories of attacks by the southern cassowary in Queensland. Memoirs of the Queensland Museum 49 (1): 335 - 338.
Kool, R. 1981. The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada. Canadian Journal of Earth Sciences 18 (4): 823 - 825.
Kowallis, B. J., Christiansen, E. H., Deino, A. L., Peterson, F., Turner, C.E., Kunk, M. J., and Obradovich, J. D., 1998, The age of the Morrison Formation. Modern Geology, v. 22, p. 235-260.
Li, R., M. G. Lockley, P. J. Makovicky, M. Matsukawa, M. A. Norell, J. D. Harris, M. Liu. 2007. Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China. Naturwissenschaften 95 (3): 185 - 191.
Lipka, T. R. 1998. The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland. In Lucas, S. G., J. I. Kirkland, J. W. Estep. Lower and Middle Cretaceous Terrestrial ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14. Albuquerque: New Mexico Museum of Natural History and Science: 229 - 234.
Long, J. A., P. Schouten. 2008. Deinonychus. Feathered Dinosaurs: The Origin of Birds. Oxford University Press: 142 - 143.
Makovicky, P. J., G. Grellet-Tinner. 2000. Association between a specimen of Deinonychus antirrhopus and theropod eggshell. In Bravo, A. M., T. Reyes. First International Symposium on Dinosaur Eggs and Babies, Isona i Conca Dellà Catalonia, Spain, 23 - 26: 123 - 128.
Manning, P. L., D. Payne, J. Pennicott, P. M. Barrett, R. A. Ennos. 2006. Dinosaur killer claws or climbing crampons? Biology Letters 2 (1): 110 - 112.
Martyniuk, M. 2016. You’re Doing It Wrong: Microraptor Tails and Mini-Wings. DinoGoss Blog.
Maxwell, W. D. 1993. Neonate dinosaur remains and dinosaur eggshell from the Lower Cretaceous Cloverly Formation, Montana. Journal of Vertebrate Paleontology 13(3, suppl.):49A
Maxwell, W. D., J. H. Ostrom. 1995. Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. Journal of Vertebrate Paleontology 15(4):707-712
Maxwell, W. D., L. M. Witmer. 1996. New Material of Deinonychus (Dinosauria, Theropoda). Journal of Vertebrate Paleontology 16 (3): 51A.
Moberly, R. M. 1960. Morrison, Cloverly, and Sykes Mountain formations, northern Bighorn basin, Wyoming and Montana: Geological Society of America Bulletin 71 (8): 1137 - 1176.
Molnar, R. E. 2001. Theropod paleopathology: a literature survey. In Mesozoic Vertebrate Life. Indiana University Press: 337 - 363.
Norell, M. A., E. S. Gaffney, L. Dingus. 1995. Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf: 126 - 130.
Norell, M. A., P. J. Makovicky. 1999. Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis. American Museum Novitates 3282: 1 - 45.
Norell, M. A., P. J. Makovicky. 2004. Dromaeosauridae. In Weishampel, D. B., P. Dodson, H. Osmólska. The Dinosauria (2nd ed.). Berkeley: University of California Press: 196 - 201.
Norell, M. A., J. M. Clark, A. H. Turner, P. J. Makovicky, R. Barsbold, T. Rowe. 2006. A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia). American Museum Novitates 3545: 1 - 51.
Nydam, R.L., and Cifelli, R.L. 2002. Lizards from the Lower Cretaceous (Aptian-Albian) Antlers and Cloverly Formations. Journal of Vertebrate Paleontology 22: 286-298.
Oreska, M. P. J., M. T. Carrano, K. M. Dzikiewicz. 2013. Vertebrate paleontology of the Cloverly Formation (Lower Cretaceous), I: faunal composition, biogeographic relationships, and sampling. Journal of Vertebrate Paleontology 33(2):264-292
Ostrom, J. H. 1969. A new theropod dinosaur from the Lower Cretaceous of Montana. Postilla 128:1-17
Ostrom, J. H. 1969. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Peabody Museum of Natural History Bulletin 30: 1 - 165.
Ostrom, J. H. 1970. Stratigraphy and paleontology of the Cloverly Formations (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana. Peabody Museum Bulletin 35:1-234
Ostrom, J. H. 1974. The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia): A Correction. Postilla, Peabody Museum of Natural History Bulletin 165: 1 - 11.
Ostrom, J. H. 1976. On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus. Breviora 439:1-21
Ostrom, J. H. 1976. Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society 8 (2): 91 - 182.
Parsons, W., K. Parsons. 2006. Morphology and size of an adult specimen of Deinonychus antirrhopus (Saurischia, Theropoda). Journal of Vertebrate Paleontology 26 (3): 109A.
Parsons, W. L., K. M. Parsons. 2009. Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda). Bulletin of the Buffalo Society of Natural Sciences: 38.
Parsons, W. L., K. M. Parsons. 2015. Morphological Variations within the Ontogeny of Deinonychus antirrhopus (Theropoda, Dromaeosauridae). PLoS ONE 10 (4): e0121476.
Pei, R., Q. Li, Q. Meng, K.-Q. Gao, M. A. Norell. 2014. A New Specimen of Microraptor (Theropoda: Dromaeosauridae) from the Lower Cretaceous of Western Liaoning, China. American Museum Novitates 3821:1-28
Prieto-Marquez, A., M. Bolortsetseg, J. R. Horner. 2012. A diminutive deinonychosaur (Dinosauria: Theropoda) from the Early Cretaceous of Oosh (Ovorkhangai, Mongolia). Alcheringa 36(1):117-136
Porfiri, J. D., J. O. Calvo, D. Santos. 2011. A new small deinonychosaur (Dinosauria: Theropoda) from the Late Cretaceous of Patagonia, Argentina. Anais da Academia Brasileira de Ciências 83(1):109-116
Redford, K. H., G. Peters. 1986. Notes on the biology and song of the red-legged seriema (Cariama cristata). Journal of Field Ornithology 47 (4): 261 - 269.
Richard L. Cifelli; Joshua E. Cohen; Brian M. Davis (2016). New tribosphenic mammals from the Mussentuchit Local Fauna (Cedar Mountain Formation, Cenomanian), Utah, USA. Palaeontologia Polonica 67: 67–81.
Roach, B. T., D. L. Brinkman. 2007. A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. Bulletin of the Peabody Museum of Natural History 48 (1): 103 - 138.
Roca-Argemi, X. and Nadon, G. C. 2003. The Buckhorn Conglomerate as the upper member of the Morrison Formation: new evidence from the type section, Cedar Mountain, Utah. Geological Society of America, Rocky Mountain Section, 55th Annual Meeting, Paper 14-1.
Rothschild, B., D. H. Tanke, T. L. Ford. 2001. Theropod stress fractures and tendon avulsions as a clue to activity. In Mesozoic Vertebrate Life. Indiana University Press: 331 - 336.
Royo-Torres, R.; Upchurch, P.; Kirkland, J.I.; DeBlieux, D.D.; Foster, J.R.; Cobos, A.; Alcalá, L. (2017). "Descendants of the Jurassic Turiasaurs from Iberia found refuge in the Early Cretaceous of western USA". Scientific Reports. 7 (1): 14311.
Sakamoto, M. 2010. Jaw biomechanics and the evolution of biting performance in theropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences 277 (1698): 3327 - 3333.
Sanders, F.; Manley, K.; Carpenter, K. (2001). Gastroliths from the Lower Cretaceous sauropod Cedarosaurus weiskopfae. In Tanke, Darren; Carpenter, Ken. Mesozoic Vertebrate Life: New Research Inspired by the Paleontology of Philip J. Currie. Indiana University Press. pp. 166–180.
Senter, P. 2006. Comparison of Forelimb Function between Deinonychus and Bambiraptor (Theropoda: Dromaeosauridae). Journal of Vertebrate Paleontology 26 (4): 897 - 906.
Shapiro, R.S., Fricke, H.C., and Fox, K. 2009. Dinosaur-bearing oncoids from ephemeral lakes of the Lower Cretaceous Cedar Mountain Formation, Utah. Palaios 2(4):51-58.
Smith-Paredes, D., D. Núñez-León, S. Soto-Acuña, J. O’Connor, J. Francisco Botelho, A. O. Vargas. 2018. Dinosaur ossification centrs in embryonic birds uncover developmental evolution of the skull. Nature Ecology & Evolution 2: 1966-1973.
Stokes, W. L. 1944, Morrison and related deposits in the Colorado Plateau. Geological Society of America Bulletin v. 55, p. 951-992.
Stokes, W.L. 1952. Lower Cretaceous in the Colorado Plateau. American Association of Petroleum Geologists v. 36: 1766-1776.
Sues, H.-D. 1980. Anatomy and relationships of a new hypsilophodontid dinosaur from the Lower Cretaceous of North America. Palaeontographica Abteilung A 169(1-3):51-72
Swanson, B. A., V. L. Santucci, S. K. Madsen, A. S. Elder, J. P. Kenworthy. 2005. Arches National Park paleontological survey. Geological Resources Division Technical Report NPS/NRGRD/GRDTR-05/01 1-36
Taylor, M. P., M. J. Wedel, R. L. Cifelli. 2011. A new sauropod dinosaur from the Lower Cretaceous Cedar Mountain Formation, Utah, USA. Acta Palaeontologica Polonica 56(1):75-98
Tennant, J. P., P. D. Mannion, P. Upchurch. 2016. Evolutionary relationships and systematics of Atoposauridae (Crocodylomorpha: Neosuchia): implications for the rise of Eusuchia. Zoological Journal of the Linnean Society 177:854-936
Therrien, F., D. M. Henderson, C. B. Huff. 2005. Bite me: biomechanical models of theropod mandibles and implications for feedinb behavior. In Carpenter, K. The Carnivorous Dinosaurs. Indianapolis: Indiana University Press: 179 - 237.
Turner, A. H., P. J. Makovicky, M. A. Norell. 2007. Feather quill knobs in the dinosaur Velociraptor. Science 317 (5845): 1721.
Turner, A. S., S. H. Hwang, M. A. Norell. 2007. A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia. American Museum Novitates 3557: 1 - 27.
S. Geological Survey. 1993. Geological Unit: Cloverly.
Varricchio, D. J. 2001. Late Cretaceous oviraptorosaur (Theropoda) dinosaurs from Montana. pp. 42–57 in D. H. Tanke and K. Carpenter (eds.), Mesozoic Vertebrate Life. Indiana University Press, Indianapolis, Indiana.
Wedel, M.J., Cifelli, R.L., and Sanders, R. K. 2000. Sauroposeidon Proteles, A new sauropod from the Early Cretaceous of Oklahoma. Journal of Vertebrate Paleontology 20:109-114.
Wedel, M. J., R. L. Cifelli. 2005. Sauroposeidon: Oklahoma’s Native Giant. Oklahoma Geology Notes 65 (2): 40 - 57.
Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press.
Wiemann, J., T.-R. Yang, M. A. Norell. 2018. Dinosaur egg colour had a single evolutionary origin. Nature 563: 555 - 558.
Williamson, T. E., S. L. Brusatte. 2014. Small theropod teeth from the Late Cretaceous of the San Juan Basin, northwestern New Mexico and their implications for understanding latest Cretaceous dinosaur evolution. PLoS ONE 9(4):e93190:1-23
Witmer, L. M., W. D. Maxwell. 1996. The skull of Deinonychus (Dinosauria: Theropoda): New insights and implications. Journal of Vertebrate Paleontology 16 (3): 73A.
White, M. A., P. R. Bell, A. G. Cook, D. G. Barnes, T. R. Tischler, B. J. Bassam, D. A. Elliot. 2015. Forearm range of motion in Australovenator wintonensis (Theropoda, Megaraptoridae). PLoS ONE 10(9):e0137709:1-20
Woodruff, D. C. 2012. A new titanosauriform from the Early Cretaceous Cloverly Formation of Montana. Cretaceous Research 36: 58 - 66.
Xu, X., Z. Zhou, X. Wang, X. Kuang, F. Zhang, X. Du. 2003. Four-winged dinosaurs from China. Nature 421 (6921): 335 - 340.
#deinonychus#deinonychus antirrhopus#raptor#dinosaur#bird#birblr#palaeoblr#dromaeosaur#cretaceous#north america#carnivore#theropod thursday#prehistory#prehistoric life#paleontology#factfile
1K notes
·
View notes
Last Seen Blogs

asphaltkissed-archived
★— 🇸🇹🇦🇷-🇧🇷🇺🇮🇸🇪🇩;

sinful-whiteknight-blog
as your knight i am your sword

aspenb
babe with the power

mychemicalblackparade
Unstoppable

blackfort
fαllıng ın love