#Mendelian transmission
Text
When a simple organism sidesteps a complex law of nature
Early people will need to have puzzled why a lady or a boy was equally doubtless at delivery. Why did they seem in a 50:50 ratio and never, say, 67:33?
We know in the present day that the reply has two elements.
How is intercourse determined at conception?
One half is that women have two X chromosomes, one acquired from the mom and the opposite from the daddy, whereas boys have an X from the mom…
View On WordPress
#Ascomycota fungi#ascospores#fumonisin#Fusarium verticillioides#Mendelian transmission#Mendel’s laws#Neurospora crassa#non-Mendelian transmission#segregation distorter
0 notes
Text
This idea that a human is a kind of natural cyborg, that we are the product of what might be called the entanglement of human nature and human invention, as philosophers as diverse as Donna Haraway and Andy Clark have proposed, is central to what I call “the New Synthesis.” Modern biology achieved its full explanatory power, its ability to account for life, its variety, and origins, thanks to the Original Synthesis, that is, the integration of Darwinian evolution with Mendelian genetics, but also with the new molecular biology that came of age in the mid-twentieth century. But if we are to explain the human mind, it is now believed by many, we need a New Synthesis, that is, we need to join biology, so understood, to the theory of cultural evolution. For it is cultural selection, and cultural modes of sharing, change, and transmission, rather than genetics or natural selection, that explain the emergence of uniquely human cognitive accomplishments, a process that, as we have already considered, probably began around one hundred thousand years ago. It is culture, layered on top of biology, that explains how we became human.
Alva Noë. 2023. The Entanglement: How Art and Philosophy Make Us What We Are. Princeton, NJ: Princeton University Press.
2 notes
·
View notes
Text
Fwd: Postdoc: UMontana_ScrippsCollege.SelfishCentromereGenomics
Begin forwarded message:
> From: [email protected]
> Subject: Postdoc: UMontana_ScrippsCollege.SelfishCentromereGenomics
> Date: 12 July 2024 at 05:25:59 BST
> To: [email protected]
>
>
>
> The Fishman (Division of Biological Sciences, University of Montana) and
> Finseth (Department of Natural Sciences, Scripps and Pitzer Colleges)
> Labs are seeking a highly motivated postdoctoral associate to join an
> NSF-funded study of the mechanisms and consequences of centromere-mediated
> meiotic drive in monkeyflowers. This project builds on robust genomic
> resources for yellow monkeyflowers to investigate the roles of satellite
> DNA, centromeric proteins, and linked genes in causing selfish chromosomal
> transmission. The postdoctoral associate's primary research responsibility
> will be genomic and epigenomic analyses of satellite DNA evolution and
> centromere formation among functionally distinct centromeres. However, the
> project is integrative and the successful candidate will be encouraged
> to participate in the other components, as well as to develop new
> research directions. Yellow monkeyflowers (Mimulus guttatus species
> complex) provide a uniquely powerful system for investigating how selfish
> centromeres violate Mendelian laws to impact natural population. Thus,
> we particularly encourage applicants interested in developing their
> own research program on genetic conflict at the interface of molecular
> and evolutionary biology. The postdoctoral scholar will also have
> opportunities to gain experience mentoring undergraduate and graduate
> students, and the option of co-teaching in an embedded CURE.
>
> The postdoc will be co-mentored by Dr. Fishman and Dr. Finseth. The
> position will be hired through the University of Montana, but the
> postdoc may choose to be physically based either in the Fishman Lab at UM
> (Missoula, Montana) or in the Finseth Lab at Scripps and Pitzer Colleges
> (Claremont, CA), with support for cross-institution travel. The University
> of Montana is an ADA/EOE/AA/Veteran's Preference Employer and has a strong
> institutional commitment to the principle of diversity in all areas.
>
>
> Position Details
> Position is full-time, 1.0 FTE, Letter of Appointment and
> includes a comprehensive and competitive benefits package
>
0 notes
Text
Understanding Chromosomal and Extrachromosomal Inheritance Patterns
The intricate world of genetics governs the transmission of traits from one generation to the next. Within this realm, two fundamental concepts play pivotal roles – Chromosomal and Extrachromosomal Inheritance. To comprehend these patterns fully, it's crucial to delve into the subtle differences between these similar-sounding terms and grasp their significance in the broader context of genetic inheritance.
Distinguishing Chromosomal and Extrachromosomal Inheritance:
Chromosomal Inheritance primarily involves the transmission of genetic information through the chromosomes, the thread-like structures that house our DNA. The nucleus of every human cell contains 23 pairs of chromosomes, each carrying a unique set of genes. During reproduction, these chromosomes are passed down from parent to offspring, determining traits ranging from eye color to susceptibility to certain diseases.
On the other hand, Extrachromosomal Inheritance refers to genetic material that exists outside the chromosomal structures. This includes mitochondrial DNA (mtDNA) and plasmids. Unlike chromosomal DNA, which is predominantly inherited from both parents, extrachromosomal elements are often maternally inherited. For instance, mitochondria, the energy-producing organelles in cells, contain their own DNA, and only the mother's egg contributes these mitochondria to the developing embryo.

Understanding the Differences:
These difference between similar terms lies in their genetic carriers. Chromosomal Inheritance involves the full set of chromosomes inherited from both parents, providing a diverse array of genetic information. In contrast, Extrachromosomal Inheritance relies on a more specific subset of genetic material, typically inherited from a single parent.
Moreover, while chromosomal traits are subject to Mendelian laws of inheritance, extrachromosomal traits often exhibit unique patterns due to their maternal transmission and distinct genetic mechanisms. This distinction is crucial in comprehending the full spectrum of genetic diversity and its implications for health and evolution.
The Bloodline Connection:
The term "bloodline" is often used metaphorically to signify the transmission of familial traits, but in the context of genetics, it is intricately linked to chromosomal inheritance. The bloodline, represented by the genetic information carried in the chromosomes, encapsulates the essence of familial connections and hereditary characteristics.
Conclusion:
In unraveling the genetic tapestry of Chromosomal and Extrachromosomal Inheritance, it is imperative to discern the subtle yet significant differences between these similar terms. Chromosomal Inheritance forms the backbone of genetic transmission, while Extrachromosomal Inheritance introduces a layer of complexity through maternally inherited genetic elements. Together, they shape the intricate mosaic of traits that define our existence and underscore the fascinating interplay of genetics in the continuum of life
0 notes
Text
Understanding Chromosomal and Extrachromosomal Inheritance Patterns
The intricate world of genetics governs the transmission of traits from one generation to the next. Within this realm, two fundamental concepts play pivotal roles – Chromosomal and Extrachromosomal Inheritance. To comprehend these patterns fully, it's crucial to delve into the subtle differences between these similar-sounding terms and grasp their significance in the broader context of genetic inheritance.
Distinguishing Chromosomal and Extrachromosomal Inheritance:
Chromosomal Inheritance primarily involves the transmission of genetic information through the chromosomes, the thread-like structures that house our DNA. The nucleus of every human cell contains 23 pairs of chromosomes, each carrying a unique set of genes. During reproduction, these chromosomes are passed down from parent to offspring, determining traits ranging from eye color to susceptibility to certain diseases.
On the other hand, Extrachromosomal Inheritance refers to genetic material that exists outside the chromosomal structures. This includes mitochondrial DNA (mtDNA) and plasmids. Unlike chromosomal DNA, which is predominantly inherited from both parents, extrachromosomal elements are often maternally inherited. For instance, mitochondria, the energy-producing organelles in cells, contain their own DNA, and only the mother's egg contributes these mitochondria to the developing embryo.

Understanding the Differences:
These difference between similar terms lies in their genetic carriers. Chromosomal Inheritance involves the full set of chromosomes inherited from both parents, providing a diverse array of genetic information. In contrast, Extrachromosomal Inheritance relies on a more specific subset of genetic material, typically inherited from a single parent.
Moreover, while chromosomal traits are subject to Mendelian laws of inheritance, extrachromosomal traits often exhibit unique patterns due to their maternal transmission and distinct genetic mechanisms. This distinction is crucial in comprehending the full spectrum of genetic diversity and its implications for health and evolution.
The Bloodline Connection:
The term "bloodline" is often used metaphorically to signify the transmission of familial traits, but in the context of genetics, it is intricately linked to chromosomal inheritance. The bloodline, represented by the genetic information carried in the chromosomes, encapsulates the essence of familial connections and hereditary characteristics.
Conclusion:
In unraveling the genetic tapestry of Chromosomal and Extrachromosomal Inheritance, it is imperative to discern the subtle yet significant differences between these similar terms. Chromosomal Inheritance forms the backbone of genetic transmission, while Extrachromosomal Inheritance introduces a layer of complexity through maternally inherited genetic elements. Together, they shape the intricate mosaic of traits that define our existence and underscore the fascinating interplay of genetics in the continuum of life
0 notes
Text
Understanding Chromosomal and Extrachromosomal Inheritance Patterns
The intricate world of genetics governs the transmission of traits from one generation to the next. Within this realm, two fundamental concepts play pivotal roles – Chromosomal and Extrachromosomal Inheritance. To comprehend these patterns fully, it's crucial to delve into the subtle differences between these similar-sounding terms and grasp their significance in the broader context of genetic inheritance.
Distinguishing Chromosomal and Extrachromosomal Inheritance:
Chromosomal Inheritance primarily involves the transmission of genetic information through the chromosomes, the thread-like structures that house our DNA. The nucleus of every human cell contains 23 pairs of chromosomes, each carrying a unique set of genes. During reproduction, these chromosomes are passed down from parent to offspring, determining traits ranging from eye color to susceptibility to certain diseases.
On the other hand, Extrachromosomal Inheritance refers to genetic material that exists outside the chromosomal structures. This includes mitochondrial DNA (mtDNA) and plasmids. Unlike chromosomal DNA, which is predominantly inherited from both parents, extrachromosomal elements are often maternally inherited. For instance, mitochondria, the energy-producing organelles in cells, contain their own DNA, and only the mother's egg contributes these mitochondria to the developing embryo.

Understanding the Differences:
These difference between similar terms lies in their genetic carriers. Chromosomal Inheritance involves the full set of chromosomes inherited from both parents, providing a diverse array of genetic information. In contrast, Extrachromosomal Inheritance relies on a more specific subset of genetic material, typically inherited from a single parent.
Moreover, while chromosomal traits are subject to Mendelian laws of inheritance, extrachromosomal traits often exhibit unique patterns due to their maternal transmission and distinct genetic mechanisms. This distinction is crucial in comprehending the full spectrum of genetic diversity and its implications for health and evolution.
The Bloodline Connection:
The term "bloodline" is often used metaphorically to signify the transmission of familial traits, but in the context of genetics, it is intricately linked to chromosomal inheritance. The bloodline, represented by the genetic information carried in the chromosomes, encapsulates the essence of familial connections and hereditary characteristics.
Conclusion:
In unraveling the genetic tapestry of Chromosomal and Extrachromosomal Inheritance, it is imperative to discern the subtle yet significant differences between these similar terms. Chromosomal Inheritance forms the backbone of genetic transmission, while Extrachromosomal Inheritance introduces a layer of complexity through maternally inherited genetic elements. Together, they shape the intricate mosaic of traits that define our existence and underscore the fascinating interplay of genetics in the continuum of life
0 notes
Text

Gene drive can be explained as a phenomenon whereby a particular gene via genetic engineering methods biases inheritance in its favour (more than the Mendelian 50:50 inheritance chance) resulting in the gene becoming more prevalent in the population over successive generations. The speed of this process is inversely correlated to the generation time of the organism, eg: it is faster in mosquitoes with a generation time of 2-4 weeks than in whales with generation time of 50 years or more.
This idea is mostly being explored as a potentially durable and cost effective strategy for controlling the transmission of deadly vector-borne diseases that effect millions of people worldwide such as malaria, dengue, Zika virus, or to eliminate herbicide or pesticide resistance. Eg: The Cas9 mediated gene drive led by Gantz et. al. in the mosquito population by introgression of parasite-resistance genes, thereby modifying the ability of the vector to transmit the pathogen.
Though the dangers of this technology cannot be overlooked. One of the ideas was a suggestion to use CRISPR gene editing technology to avert extinction of endangered wildlife by spreading a fertility reducing gene in the other animals competing against them for resources. This with time turned from an innovative idea to a perilious one, as demonstrated in a CRISPR-Cas9 gene drive targetted at genes that control the differentiation of the two sexes of the Anopheles gambiae mosquitoes. Leaving the males unaffected it decreased the fertility of the female mosquitoes. The gene rapidly spread reaching 100% prevalence within 7-11 generation to the point of total population collapse despite various Cas9-resistant genes arising in each generation.
With just a few of these engineered organisms our ecosystem could be altered, irrevocably.
References: 1) Gantz, V. M., Jasinskiene, N., Tatarenkova, O., Fazekas, A., Macias, V. M., Bier, E., & James, A. A. (2015). Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proceedings of the National Academy of Sciences, 112(49), E6736. https://doi.org/10.1073/pnas.1521077112
2) Kyrou, K., Hammond, A., Galizi, R. et al. A CRISPR–Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat Biotechnol 36, 1062–1066 (2018). https://doi.org/10.1038/nbt.4245
3) Emerson, C., James, S., Littler, K., & Randazzo, F. (Fil). (2017). Principles for gene drive research. Science, 358(6367), 1135. https://doi.org/10.1126/science.aap9026
4) Alphey, L. S., Crisanti, A., Randazzo, F. (Fil), & Akbari, O. S. (2020). Opinion: Standardizing the definition of gene drive. Proceedings of the National Academy of Sciences, 117(49), 30864. https://doi.org/10.1073/pnas.2020417117
#biotechnology#biology#molecularbiology#bobthebiotechquer#dna#molbio#studyblr#genedrive#ecosystem#crispr#editing#genetics#malaria#dengue#mosquito
64 notes
·
View notes
Text
Mendelian experiment
reproduction in living organisms is done to facilitate the continuity of life. The progeny produced, often resemble their parents in most of the characteristics. This passage or transmission of characters from parents to their offsprings is called Heredity
2 notes
·
View notes
Note
I found your blog through that post on DNA being utter chaos. I hope I’m not being annoying, but I start college this fall and I’ll be majoring in genetics (my dream ever since I was little). I’m really nervous though; I bought all these genetics-based books and I did labs at Cold Spring Harbor Labs for six summers. I also wrote a paper on transposons causing cancer last year. I promise I’m not totally new to this (but still pretty new),but would you maybe consider giving me some advice/tips?
I love that you asked this! It sounds to me like you’re super prepared already! Remember, the classes in college are for you to learn--nobody should expect you to come in already knowing everything about genetics. In the intro genetics course that I teach, we hardly even talk about transposons (not because they’re not cool, but just because there’s only so much time), so you’re already ahead of my students there! I think you’re coming it already at a really great place, and excited to learn more (which is the most important part, really).
But, a few pieces of advice for studying genetics (which are going to be tilted towards transmission genetics rather than molecular genetics, because that’s the part I teach):
- Be really comfortable with basic probability. Transmission genetics--things like Mendelian genetics, Punnet Squares, and more complicated things like recombination mapping (if you don’t know about that yet, don’t worry) are all probabalistic. You don’t need complex probability skills, but it helps if you’re really comfortable with thinking about how you can predict something based on probability, the probability that two events will both happen (multiply them) or that either will happen (add them), etc. If you’re struggling to follow the probability stuff, it’ll be a lot harder to learn how to relate it to the biology.
- Genetics is largely a problem-based discipline, especially the Mendelian/transmission stuff. You’ll learn best if you do a lot of practice problems as well as just studying your notes. Hopefully your class will include a lot of opportunities to do that, but if not you can seek them out on your own; your textbook will definitely have some at the ends of chapters, you can write some and exchange them with friends, etc.
- More generally, as far as studying, don’t just go back over your notes. Do practice problems. Write practice problems. Re-write your notes. Make notecards. Draw chromosomes during replication and label every part that you need to know. Do as much of it from memory as you can, then check your work. Retrieving information is a different thing than just re-reading it; you want to practice retrieving specifically, because that’s what you have to do on exams. The more often you have to remember a specific piece of information at home, the more likely that you’ll be able to later.
- Study often, not just before exams. Lectures are going to build on themselves, so if you can’t remember the previous week it’s going to be hard to really follow the current one. And try to keep up with the vocab! The professor isn’t teaching vocab to be annoying, but because you need to know the words in order to talk about the biology (every time I try to reduce vocab in my course, it ends up actually being more confusing for students, because they don’t have the terms to talk clearly about what’s going on). Studying often will help keep you from being overwhelmed.
- That being said, for many (not all!) students there are some times when it just seems like there is too much information to learn. More than their head can hold. What that usually means is that you’re trying to memorize things as separate, individual pieces of information. There is actually a limit to how many pieces of individual information you can hold at once. If that’s the case, what it means is that you need a better way of organizing it--linking concepts/vocab/facts up together so that it’s not so many little pieces to remember. In education circles, it’s called chunking information. A good professor will be helping you do this, but you can also do some of it yourself. Things like making concept maps can be really helpful in getting you to think about how things connect.
- The biggest one: if you need help, ask for it. If your classes aren’t very large, then the professor is probably your first go-to. They should have “office hours” listed in the syllabus; those are times when they are in their office, ready for students to drop in with questions. Go to them! Go if you’re confused and need help, or go if you were really excited about something in class and want to ask more about it. If you’re at a larger school and the classes are big, then it’s more likely that your TA will be the first go-to, but that’s fine too. They’ll probably also have office hours. But, whoever it is you go to, ask for help if you need it! And ask early. Don’t wait until you’re lost on three weeks of material or you think you’re about to fail a test. If there’s something from a lecture that you don’t understand, ask for help that week instead of waiting. That is absolutely the most important piece of advice I can give any college student.
- Oh, and the other main piece of advice: Go to class. Go to class. Please, please, go to class. Reading the textbook, doing the homework, or even getting notes from a friend is not a substitute. In every class I’ve taught, there’s been a signficant linear relationship between attendance and grades. Please go to class.
Wow, apparently I’ve been saving a lot of this up just waiting for someone to ask. I hope this is more encouraging/helpful than daunting. I really think that you’re going to do great! And I’m so excited for you--genetics is a really, really awesome discipline!
2 notes
·
View notes
Text
Super-Mendelian inheritance mediated by CRISPR–Cas9 in the female mouse germline
A gene drive biases the transmission of one of the two copies of a gene such that it is inherited more frequently than by random segregation. Highly efficient gene drive systems have recently been developed in insects, which leverage the sequence-targeted DNA cleavage activity of CRISPR–Cas9 and endogenous homology-directed repair mechanisms to convert heterozygous genotypes to homozygosity1,2,3,4. If implemented in laboratory rodents, similar systems would enable the rapid assembly of currently impractical genotypes that involve multiple homozygous genes (for example, to model multigenic human diseases).
To our knowledge, however, such a system has not yet been demonstrated in mammals. Here we use an active genetic element that encodes a guide RNA, which is embedded in the mouse tyrosinase (Tyr) gene, to evaluate whether targeted gene conversion can occur when CRISPR–Cas9 is active in the early embryo or in the developing germline. Although Cas9 efficiently induces double-stranded DNA breaks in the early embryo and male germline, these breaks are not corrected by homology-directed repair. By contrast, Cas9 expression limited to the female germline induces double-stranded breaks that are corrected by homology-directed repair, which copies the active genetic element from the donor to the receiver chromosome and increases its rate of inheritance in the next generation.
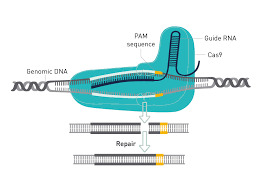
These results demonstrate the feasibility of CRISPR–Cas9-mediated systems that bias inheritance of desired alleles in mice and that have the potential to transform the use of rodent models in basic and biomedical research.
9 notes
·
View notes
Text
The Value of W, or, Interdisciplinary Engagements on Culture
OCTOBER 31, 2018
LAST SPRING, I attended a conference in New Mexico featuring evolutionary biologists working on a new research program they have been calling the “extended evolutionary synthesis” (EES). The program aims to go beyond the so-called “modern synthesis” of the mid-20th century, which joined Darwinism to Mendelian genetics, whose mathematical formulations could be simply and straightforwardly expressed. Biologists involved in the EES have been calling for a broader and less reductive view of evolution, unrestricted to Mendelian genes. In particular, they have been addressing the modern synthesis’s paucity of information about developmental biology. These EES revisionists are interested in feedbacks: in how developmental processes, along with ecological and even cultural ones, feed back into one another, into genetic and other forms of inheritance, and therefore into evolution. While the modern synthesis proposes that epigenetic, developmental, ecological, and cultural processes are all products of evolution, the EES claims they are causes as well as products.
The roots of this movement extend back to the early 1970s, to the work of Richard Lewontin at Harvard, and Luigi Luca Cavalli-Sforza and Marcus Feldman at Stanford, among others. But the reigning, reductive neo-Darwinist paradigm — in other words, the modern synthesis — remains well entrenched, and its defenders staunch in its support. Only in the last 25 years or so has the more expansive vision of the EES slowly begun — against much resistance — to establish itself in mainstream biology.
As part of this development, EES biologists have been increasingly interested in culture, among other forms of transformation and transmission, and so have welcomed the input of humanists, including philosophers and historians of science like me, whose job it is to study and understand culture. Taking part in their conversations has in turn informed my own work in the history of evolutionary theory.
Still, I experienced a moment of comical culture shock at the recent meeting I attended. A biologist wrote an equation on the whiteboard in which one of the variables was a “w.” He then circled the w, explaining that it represented “culture,” and pointed out that under certain conditions, the value of “w” would tend toward zero, while under other conditions it would tend toward 100. “Perhaps,” I thought, “we don’t mean quite the same thing by ‘culture.’” To a humanist, or anyway to this one, “culture” is an abstract noun encompassing many things of many kinds: processes, objects, habits, beliefs both explicit and implicit. It seems a category mistake to think that we can represent such a welter by a single variable, or that the whole jumble could act as a discrete thing having a single quantifiable effect on some other discrete thing. Could we say, for example, that in a given society, “culture” influences “politics” by some quantifiable amount x? Could we say that “the arts” has a y-percent effect on birth rate or life expectancy?
As it turned out, I had somewhat misunderstood the situation. When I expressed a certain dubiousness about representing “culture” with a single variable, an EES biologist explained to me that the variables standing for “culture” in biologists’ mathematical models are not meant to denote the entire Gestalt, but rather quantifiable bits of culture: a single behavior, for example, that might be taught, learned, transmitted, or counted, and whose effects on survival and reproduction can be measured and modeled. Perhaps these individual culture variables might in principle add up to a single, overarching W, but for the moment, no one claims to be able to make that summation. For now, we can simply use the little w’s to build discrete cultural bits or forms into an evolutionary model. This seems to me more credible, but it still assumes that we can meaningfully represent cultural forms as quantifiable bits, and that this will add more to our understanding of the role of cultural forms in evolutionary processes than simply trying to describe this role in qualitative terms. I can’t help wondering if that’s a sound assumption.
Of course I’m by no means the first to raise the question, nor indeed have such objections been confined to humanists. Lewontin himself, together with the historian Joseph Fracchia, argued in a 1999 paper against the idea of cultural evolution. They wondered whether conceptualizing entities like “the idea of monotheism” as “cultural units” begged crucial questions — for example, how can we count up these units in a population, and what are their laws of inheritance and variation? Fracchia and Lewontin maintained that there could be no such general laws because cultural phenomena, unlike atoms and molecules, differ from one another in their properties and dynamics of transmission and change. “There is no one transhistorical law or generality,” they contended, “that can explain the dynamics of all historical change.” [1] Marcus Feldman disagreed, albeit not specifically with regard to the existence of general laws explaining the dynamics of all historical change; rather, he defended the notion of “observable units of culture,” which he did not associate with grand organizing ideas such as monotheism. An example of an observable cultural unit for Feldman is a behavior or custom that follows statistical rules of transmission, and that can therefore be a legitimate object of mathematical study. [2]
The biologist with the “w” variable and I were thus reenacting an intellectual confrontation that has been going on for decades. As is often the case in longstanding debates, we actually agree on the essentials: that nature and culture are at bottom made of the same stuff — in fact, of one another — which no humanistic or scientific inquiry can legitimately disregard. Evolutionary theory must encompass cultural processes just as human history must encompass biological ones. But, despite our deep accord, this biologist and I are thinking incommensurately about methods, about how to put our two fields into communication. His method is mathematical modeling, and mine is thick description. These are diametrically opposite in trajectory, one abstractive and reductive, the other concretizing and expansive. While I understand and admire these biologists’ conclusions, I keep wondering: Why these methods? Why mathematical modeling? By which I mean, what function do biologists intend their mathematical models to serve? Are they meant to prove claims about evolution? Or rather to express, represent, or advocate certain interpretive views of evolutionary processes? If the latter, why choose this particular means of expression, representation, advocacy? These will surely seem naïve questions to any biologist reading this. But I have learned from teaching college freshman and sophomores that naïve questions from untrained newcomers can be the hardest and most useful, which emboldens me to ask mine.
¤
Kevin Laland, author of Darwin’s Unfinished Symphony, was an organizer of the conference I attended and is a leader of the Extended Evolutionary Synthesis research program. Reading his important and heartfelt book, the to-date summary of a groundbreaking career, I had similar feelings as I did at the conference. Uppermost among these is heated agreement: Laland’s essential tenets seem to me profoundly right, some indeed incontrovertible. These include the precept that cultural practices — in particular teaching, imitating, and copying — are causes as well as results of evolution; that in mammals and especially humans, such cultural practices have accelerated evolutionary development by constantly creating “new selection regimes” in a process that Laland, citing evolutionary biologist Allan Wilson, calls “cultural drive”; and that, accordingly, in humans especially, there has been a “gene-culture coevolutionary dynamic.” The first of these — that cultural practices are causes as well as results of evolution — seems to me incontrovertible, but more like a first principle than like an empirical result. Cultural practices must be causes as well as results of evolution because any result of the evolutionary process becomes a feature of the world of causes shaping the continuation of that process.
The other principal tenets — such as “cultural drive” and “gene-culture co-evolution” — are not quite first principles, but they seem to me ways of understanding how the feedback-loop of evolution encompasses cultural forms. To express these ways of understanding in the language of mathematical modeling seems fine, if one likes to do that, but no more definitive than expressing them in words. This is because a mathematical model, like a verbal description, contains many layers of interpretation. This is not a criticism: interpretation is essential to (and ineradicable from) any attempt to understand the world. But insofar as a mathematical model is taken to prove rather than to argue or represent, that’s where I think it can mislead.
Laland has devoted his career to pioneering work against reductive, simplistic, and dogmatic accounts of evolution, building brick by brick a sound case for the richer and more complex vision of the EES. Darwin’s Unfinished Symphony is a record of his resounding success. But while he has been constructing this revisionist scientific theory, he has often supported it by traditional methods. An example is his game-theoretic tournament to study social learning. After offering several examples of social learning in animals — such as Japanese macaques who learn from one innovative macaque to wash their sweet potatoes before eating them, and fish who learn from one another where the rich feeding patches are located — Laland asks what might be the best “social learning strategy.” He explains that the “traditional means to address such questions is to build mathematical models using, for instance, the methods of evolutionary game theory.”
Game theory became a standard model in evolutionary biology in the early 1970s with the work, notably, of the British theoretical evolutionary biologists W. D. Hamilton and John Maynard Smith, along with the population geneticist George R. Price. Hamilton, Price, and Maynard Smith developed a game-theoretic approach to modeling the behaviors of organisms in the struggle for survival. Their work was foundational to the neo-Darwinist, gene-centric program that Laland has devoted his career to challenging. In this gene-centric view, all higher-order entities — individual organisms, their behaviors and interactions — are epiphenomenal, controlled by and reducible to genes, so that any apparent agency or intention on the part of an organism is illusory. Organisms survive if they happen to achieve an optimal state of genetic affairs, one that maximizes some function for greater reproductive success. They die out when they fail to do so. Maynard Smith accordingly emphasized that his technical definition of “strategy” was strictly behaviorist. “Nothing,” he maintained, “is implied about intention.” A strategy was merely “a behavioural phenotype,” in other words, “a specification of what an individual will do [in a given situation].” [3] These “strategies,” therefore, involved no ascription of internal agency, but merely outward observations of behavior. Neither observed behaviors nor any other macrolevel phenomenon could play a causal role in evolution according to this school of thought.
Maynard Smith’s approach has inspired the most reductive of neo-Darwinists. For example, Richard Dawkins has adapted it to his own theory of gene functioning, emphasizing that the “strategies” in question are behaviorally defined and do not require the ascription of consciousness, let alone agency, to the strategic agent. Dawkins indeed refers to “unconscious strategists,” the deliberate oxymoron encouraging the reader to accept these apparent ascriptions of agency to genes as radical denials of any such agency. [4] Neither behaviors, nor agency, nor consciousness, nor culture operates causally at any level of Dawkins’s picture; all reduces to just gene functioning.
Game-theoretic modeling has been a hallmark of neo-Darwinist reductionism and, specifically, of the denial of any kind of evolutionary agency to the evolving organism. But in Darwin’s Unfinished Symphony, Laland describes how he and his collaborators used game theory in an innovative way, to design a virtual world in which they hosted a tournament. The game involved virtual “organisms” or “agents” engaging in a hundred “behavior patterns,” with varying rates of success resulting in greater or lesser “fitness” (i.e., survival and reproduction). The game also included three different “moves” — “innovate,” “observe,” and “exploit” — representing different phases of asocial or social learning. More than a hundred people of various ages and backgrounds took part in the game. Unlike in Maynard Smith’s applications of game theory to evolution, Laland and his collaborators were not looking for an optimum in the form of a single function or property to be maximized. They did not pre-judge what had to happen in order for an organism to win the competition. Rather, they set the competitors loose and waited to see who would triumph. The winning strategy was an unpredictable, complex mix of behaviors, although it did represent an overall optimum solution composed of behavioral bits.
Analyzing the winning strategy, Laland concludes that observing and copying are tremendously valuable, much more so than innovating on one’s own except “in extreme environments that change at extraordinarily high rates,” which must be rare in nature. The conclusion is persuasive, but the tournament seems to me more a way of expressing than of proving this point: the virtual agents and their behaviors and strategies of course constitute an interpretive representation of natural processes. They are not drawn in pastels or composed in prose, but the fact that they are programmed on a computer makes them no less a representation.
¤
To elaborate further, consider an experiment Laland describes, performed with his postdoctoral student Hannah Lewis. Laland explains that to model the effects of high-fidelity transmission of information on the longevity of cultural forms or “traits” in a population, he and Lewis “assumed that there are a fixed number of traits that could appear within a group through novel inventions and that are independent of any other traits within a culture. We called these novel inventions ‘cultural seed traits.’ Then, one of four possible events could occur”: a new seed trait could be acquired by novel invention; two traits could be combined to produce a new one; one trait could be modified; or a trait could be lost.
This model, in its relation to real cultural forms, seems to me the equivalent of a Cubist painting. Cultural “traits” that are independent of one another occur no more often in nature than young ladies with perfectly geometrical features distributed all on one side of their two-dimensional heads. Likewise for the separate and distinct occurrence of novel invention, combination, modification, or loss of cultural forms. These processes travel in the real world as aspects of a single organic entity and not as separate blocks. Of course, I’m not opposed to representing cultural forms in these Cubist terms any more than I’m opposed to Picasso’s portraits of Dora Maar. Representations should, though, declare themselves as such.
Mathematical models are interpretative from the get-go. Again, let me be clear that I think that’s fine — indeed, inevitable — because interpretation is ineradicable from any attempt to understand the world. Indeed, some scientists describe their use of mathematical models in these very terms. The theoretical physicist Murray Gell-Mann warned that we must be careful, regarding models, “not to take them too seriously but rather to use them as prostheses for the imagination, as sources of inspiration, as acknowledged metaphors. In that way I think they can be valuable.” [5] Feldman, who pointed me to Gell-Mann’s characterization of models as “prostheses of the imagination,” added that “insofar as the model assists in the interpretation, then it has value.” [6] On another occasion, Feldman told an interviewer, “[p]eople who make models for a living like I do don’t actually believe they’re describing reality. We aren’t saying that our model is more probable than another model; we’re saying it exposes what is possible.” [7]
I have no trouble believing in mathematical modeling as a powerful form of metaphor, representation of the possible, or prosthesis for the imagination. But mathematical modeling does have a distinctive feature that sets it apart from other interpretive modes: notwithstanding Gell-Mann and Feldman, it tends to disguise itself as proof rather than representation. Would it be possible for it to come right out of the positivist closet? To put my point another way, culture plays as crucial a role in evolutionary theory as it does in evolution. Culture plays as crucial a role in science as it does in nature. Wouldn’t a scientific method that unapologetically declared itself as interpretive and representational be in keeping with Laland’s revolutionary program to write cultural forms into evolutionary theory?
Mathematical modeling, like any mode of interpretive analysis, also has its limitations and pitfalls. For example, it brings a tendency I’ll call “either/or-ism”: a tendency to represent as separate and discrete, the better to count them, things that are in fact mixed and blended. Darwin’s Unfinished Symphony lists as discrete alternatives, for example, animals learning innovations socially from one another versus inventing them independently; the cultural drive hypothesis operating through natural selection on social learning proficiency versus social learning incidence; humans being more accomplished than other primates due to “chance factors” or because of a “trait or combination of traits that were uniquely possessed by our ancestors”; that high-fidelity transmission of information might have been achieved by our ancestors through language or alternatively through teaching; learning a skill such as stone-knapping to make a cutting tool by reverse-engineering from a finished sharpened flake, or else by imitation, or else by various forms of non-verbal teaching; or else by verbal teaching; and young individuals acquiring skills either asocially by trial and error, or else socially by copying, or else socially by being taught by a tutor “at some cost to the tutor.” In each of these cases, “both, and” seems more plausible to me than “either, or.” (Additionally, in the last case, must teaching involve a cost to the tutor? In my experience, teaching is often a win-win process, a non-zero-sum game, in which the teacher learns at least as much as the pupil, rather than a donation by the teacher to the pupil. Perhaps the sort of teaching that humans do is qualitatively different from the sorts that other animals do: a teacher macaque might not derive the same intellectual benefits from teaching to compensate for the loss of time that could be spent eating or reproducing. But I wonder if that’s necessarily true in all cases of nonhuman teachers.)
Yet Laland’s conclusions are extremely persuasive. Their persuasiveness overwhelms my failure to believe in a proof-value for the mathematical models. He concludes that natural selection favors those who copy others efficiently, strategically, and accurately; that nonhuman species lack cumulative cultures because of their “low-fidelity copying mechanisms”; that teaching evolves where the benefits outweigh the costs; and that language first evolved to teach close kin. I can believe in these conclusions, not as proven by the tournament-experiment, or the cultural-trait-transmission model, or the other mathematical models, but as interpretively, argumentatively presented by these models. I think this is because Laland’s conclusions are based on the kind of profound knowledge that comes only from a wealth of direct experience and — yes — keen, richly informed interpretation. Alongside the mathematical models are descriptions drawn from experiments and observations, some extending over decades.
For example, Laland describes several series of experiments designed to show that fish can learn from one another, and to investigate how and under what conditions they do so. In one set of experiments, Laland and his students and collaborators trained guppies to take certain routes to find rich food supplies, then observed other untrained guppies, in various conditions, learn from their trained fellows. In one variation, the experimenters trained the demonstrator fish to swim directly up narrow vertical tubes to reach their meal; this was a highly esoteric skill that no fish figured out on its own, without training, but the guppies did readily learn it from one another. In another series of experiments, the experimenters offered certain stickleback fish rich feeding patches and others poor ones, while observer fish watched from a distance; the humans then observed the observer fish to see whether and what they learned.
Such experiments, Laland reports, have established certain social tendencies in fish. These include “a tendency to adopt the majority behavior,” “copying the behavior of others when uncertain,” and “disproportionately attending to the behavior of groups.” Such social tendencies, once established, must surely enter into any legitimate evolutionary picture of fish. More generally, the principle that many animals are social, and that their sociality necessarily plays a role in the evolutionary process, has the retrospective obviousness of all grand, organizing ideas once stated, a most notable example being the idea of natural selection itself, whose retrospective obviousness led T. H. Huxley, upon reading the On the Origin of Species, to figuratively smack his forehead, exclaiming: “How extremely stupid of me not to have thought of that!” [8] Such grand, organizing ideas, which create conceptual sea-changes that render them retrospectively (but only retrospectively) obvious, can emerge only from richly informed interpretative analysis.
Darwin’s own method was explicitly so. He described natural history as a form of deeply interpretive historical scholarship. The geological record, he said, was a collection of fragments of the most recent volume of “a history of the world imperfectly kept, and written in a changing dialect.” He urged people to join him in considering natural history in these terms: to “regard every production of nature as one which has had a history” to be pieced together by interpretation of scant evidence. Darwin promised that this approach would be its own reward: “[W]hen we thus view each organic being, how far more interesting, I speak from experience, will the study of natural history become!” [9] Laland’s evolutionary science, as portrayed in Darwin’s Unfinished Symphony, might as well come right out and declare itself as such: it is precisely that “far more interesting” study.
¤
Jessica Riskin is a history professor at Stanford University, where she teaches courses in European intellectual and cultural history and the history of science. She is the author, most recently, of The Restless Clock: A History of the Centuries-Long Argument Over What Makes Living Things Tick (2016).
¤
[1] Joseph Fracchia and R. C. Lewontin, “Does Culture Evolve,” in History and Theory Vol. 38, No. 4, Theme Issue 38: The Return of Science: Evolutionary Ideas and History (Dec., 1999), pp. 52–78, on pp. 60, 72.
[2] Marcus W. Feldman, “Dissent with Modification: Cultural Evolution and Social Niche Construction,” in Melissa J. Brown, ed., Explaining Culture Scientifically (Seattle: University of Washington Press, 2008), Ch. 3, on p. 58.
[3] John Maynard Smith, Evolution and the Theory of Games (Cambridge: Cambridge University Press, 1982), 5, 10.
[4] Richard Dawkins, The Selfish Gene (1976), 30th anniversary ed. (Oxford: Oxford University Press, 2006), 229.
[5] Murray Gell-Mann, “Plectics,” in John Brockman, ed., Third Culture: Beyond the Scientific Revolution (New York: Touchstone, 1995), Ch. 19, on p. 324.
[6] Marcus Feldman, in conversation, August 2018.
[7] Feldman, quoted in Elizabeth Svoboda, “Finding the Actions that Alter Evolution,” in Quanta Magazine, Jaunary 5, 2017, https://www.quantamagazine.org/culture-meets-evolution-the-marcus-feldman-qa-20170105/.
[8] Thomas Henry Huxley, “On the Reception of The Origin of Species” (1887), in The Life and Letters of Charles Darwin, edited by Francis Darwin (New York: D. Appleton, 1896), 1:533–58, on p. 551.
[9] Charles Darwin, On the Origin of Species by means of natural selection, or the preservation of favoured races in the struggle for life (London: John Murray, 1859 [1st ed.]), 310–311, 485–486.
Source: https://lareviewofbooks.org/article/the-value-of-w-or-interdisciplinary-engagements-on-culture/
1 note
·
View note
Text
In a weird twist, Sperm Have Been Caught Poisoning Other Sperm to Get Ahead in Mice
https://sciencespies.com/nature/in-a-weird-twist-sperm-have-been-caught-poisoning-other-sperm-to-get-ahead-in-mice/
In a weird twist, Sperm Have Been Caught Poisoning Other Sperm to Get Ahead in Mice
When sperm race, it’s for keeps. So it comes as little surprise that in some species, the competition over who reaches the egg first can get a little dirty.
A variant in mice genes has been found to give sperm that possess it a clear advantage by poisoning its peers while they’re still in development, robbing them of their ability to efficiently sniff their way towards the egg.
In a rather karmic twist, a race consisting only of would-be assassins would be a complete disaster, with researchers finding the genetic variant risks overdosing on its own killer cocktail unless the race is balanced out by its victims.
Geneticists from the Max Planck Institute for Molecular Genetics in Berlin uncovered this rather unique ‘cheat code’ in mice sperm while investigating the mechanisms male sex cells use to direct their way through the female reproductive system.
“Our data highlight the fact that sperm cells are ruthless competitors,” says institute director Bernhard Herrmann.
“Genetic differences can give individual sperm an advantage in the race for life, thus promoting the transmission of particular gene variants to the next generation.”
They found a Rho protein switch called RAC1 plays an integral role in keeping sperm on the straight and narrow. Mess this regulator up and sperm will stagger about like it’s heading home after closing time on cheap-drinks Tuesday.
But apparently evolution has this all worked out. According to the researchers, a variation in the coding of sequences on chromosome 17 seems to do just this, churning out a product that throws a spanner in the works of RAC1.
This region – called a t-variant – isn’t new to science. In fact this stretch of DNA has stood out as an oddity in Mendelian genetics for close to a century.
Mice heterozygous for the trait (having one t-variant chromosome 17, and a partner chromosome with ‘normal’ coding) don’t father the expected 50-50 ratio of offspring you’d expect. The odds of one of their offspring being born without the t-variant, in fact, are one in a hundred.
Yet if they happen to be homozygous for it – with both versions of chromosome 17 containing this aberrant coding – then they can kiss fatherhood goodbye. They’re completely sterile.
With all of this in mind, the researchers have teased apart exactly what’s going on inside those tiny mice testicles by genotyping individual sperm and assessing their motility patterns.
Early in gamete production, inside sperm precursor cells that contain both the t-variant chromosome and a more normal version, the toxic t-coding interferes with the development of RAC1, effectively disabling it.
Once the precursor cells eventually split into their sperm forms, however, they undergo the process of meiosis, divvying up the chromosomes so each sperm only has one of each pair.
This means some sperm now have a t-variant chromosome, and others don’t. Here’s the truly clever part – the t-variants also produce their own remedy, rescuing RAC1 from harm by expressing a special regulating protein.
“Imagine a marathon, in which all participants get poisoned drinking water, but some runners also take an antidote,” says Herrmann.
That antidote works well in small enough doses. Unfortunately, an abundance of RAC1 is just as bad as a shortfall. In a marathon packed with poisoners all churning out antidotes, the racers will soon be burdened by excess RAC1.
“The competitiveness of individual sperm seems to depend on an optimal level of active RAC1; both reduced or excessive RAC1 activity interferes with effective forward movement,” says the study’s lead author Alexandra Amaral.
It’s the first time experiments have demonstrated exactly how heterozygous t-variant mice gain an advantage, while affirming the biochemistry of RAC1 in mammalian sperm navigation.
Being observed in mice, the research only has limited relevance to human reproduction. But the more we understand about diverse models of reproductive chemistry across the animal kingdom, the better we understand how ours evolved.
This research was published in PLOS One.
#Nature
0 notes
Text
experimental validation: chromosomal theory of inheritance
confirming gene inheritance via physical chromosomal movement
criteria that the chromosome theory of inheritance must meet for validation:
1. all chromosome inheritance patterns must match gene trait inheritance patterns in every detail
2. a specific chromosome suspected to contain a corresponding genetic trait must be present in an individual with relevant trait
nomenclature for chromosomal gene inheritance:
lowercase indicates recessive mutant phenotype | uppercase indicates dominant mutant phenotype
+ signage signifies wild-type phenotype individual
criterion 1. matching characteristics of meiosis to chromosomal transmission
two gene copies in each organism | two copies of each chromosome in each cell nucleus
gene from parent is conserved in offspring | chromosome appears unchanged when passed from parent to child
gene alleles segregate to different haploid gametes | homolog chromosomes separate to different cells during meiosis I
each gene assorts independently from any others | homologs segregate independently by pair and undergo intra-chromosomal recombination during meiosis I
parental gametic genes pair at random | fertilization of zygote creates random combination of maternal and paternal haploid chromosome karyotypes
criterion 1. recombinant crossing over in meiosis I fits with mendelian laws
law of segregation: barring abnormal nondisjunction events, crossing over does not affect gametes’ reception of (1) gene allele each
law of independent assortment: without recombination in reductive meiosis I division, sister chromatids would not be physically able to assort independently during meiosis II conservative division
law of independent assortment: recombination retains independent assortment of two genes that are located on the same chromosome from one another
criterion 2. specific trait appears in tandem with presence of particular chromosome within individual’s karyotype
drosophila eye-color trait phenotype corresponds with presence of X(w) or X(w+) chromosome
determined by F1 offspring phenotypes and F2 breeding ratios
eye-color gene presence on drosophila X chromosome evidenced by crisscross inheritance - males get eye-color from maternal parent, the source of single male X chromosome
X-linkage - gene is located on the X chromosome
males are hemizygous - only one copy of gene present | in genetics problems, treat like heterozygote with some considerations in mind
criterion 2. nondisjunction events show that presence of chromosome corresponds with appearance of phenotype
large-scale drosophila eye-color breeding reveals extremely rare “switch” in X-linked phenotypes
actually indicative of improper X chromosome transfer in meiotic nondisjunction event, providing strong evidence that X chromosome presence is necessary for phenotypic appearance
0 notes
Text
Fwd: Graduate position: UKiel.SelfishChromosomes
Begin forwarded message: > From: [email protected] > Subject: Graduate position: UKiel.SelfishChromosomes > Date: 16 June 2020 at 07:10:14 BST > To: [email protected] > > > *PhD position on **the evolutionary maintenance of selfish accessory > chromosomes*** > > A PhD position is available for three years to study the *mechanism of > the evolutionary maintenance of accessory chromosomes in a fungal plant > pathogen *with the Max Planck research group “Environmental Genomics” > headed by Prof. Dr. Eva H. Stukenbrock and supervised by Dr. Michael > Habig. The Max Planck group is affiliated with the *Max Planck Institute > for Evolutionary Biology* in Plön and the *Christian-Albrechts > University of Kiel* in the North of Germany. The position will also be > associated with the Kiel Evolution Center, the Kiel Plant Center and the > International Max Planck Research School for Evolutionary Biology – > providing a unique scientific environment. The position is compensated > at 65% of TV-L 13 and candidates are expected to complete a PhD. > > *Background*** > > Selfish genetic elements are widespread in all groups of organisms. > These elements are likely to have shaped the evolution of diverse > biological systems, including genome structure, sex determination, and > meiosis. To date, surprisingly little is known about the exact processes > that underlie the spread of these elements. This project aims at > understanding the transmission mechanism and its evolutionary > consequences of selfish accessory chromosomes in a fungal pathogen with > a particular focus on the role of histone modifications. > > Recently, we could show that the accessory chromosome of the > commercially important wheat pathogen /Zymoseptoria tritici/ show a > chromosome drive, i.e. are inherited to more progeny than expected by > Mendelian segregation. In addition these chromosomes show a negative > fitness effect and could therefore be considered selfish elements. The > mechanism of this chromosome drive is however unknown – but recent data > suggest that histone modifications might be involved.This project aims > at understanding this previously unknown aspect of histone modifications > and its exploitation by selfish chromosomes using the model organism /Z. > tritici/.It will involve the establishment of in vitro crosses and the > identification of the genetic and epigentic traits responsible for the > chromosome drive. > > *Expectations and Requirements:* > > We seek a highly motivated candidate who wants to actively drive a > research project. The candidate must have university degree (Master or > equivalent) in biology or a related field. High competence in > experimental design, and English writing is a must. Ideally, the > candidate has comprehensive experience in molecular genetics, work with > plant pathogens or performance of evolution experiments. The selected > candidate will have the opportunity to collaborate in an > interdisciplinary team of biologists (molecular biologists, evolutionary > biologists and population geneticists) as well as to pursue unique > research in the field of chromosome biology. > > Kiel University aims at a higher proportion of women in research and > education, and, therefore, specifically encourages qualified female > scientists to apply. Female scientists will be preferentially considered > in case of equivalent qualification, competence and achievements. > > Kiel University specifically supports employment of severely handicapped > people. Therefore, severely handicapped applicants will be > preferentially considered in case of suitable qualification. > > Kiel University specifically welcomes application from people with > migration background. > > *Application* > > Applications should include a motivation letter (max. 2 pages long), CV, > publication list, names and contact details of two referees (who are > familiar with the applicant’s work), and copies of certificates. > Deadline for applications is 15. July 2020. Applications should be sent to: > > *Dr. Michael Habig, Botanisches Institut, > Christian-Albrechts-Universität zu Kiel, Olshausenstr. 40, 24118 Kiel* > > *or _preferentially as a single-PDF-document_ by e-mail to: > [email protected]* > > We explicitly do not require application photos and therefore these > shall be omitted. > > Further information can be obtained from Dr. Michael Habig > ([email protected] ). > > Please also check: > > https://ift.tt/37zkrvo > > https://ift.tt/35psMkC > > https://ift.tt/2Gt6YJJ > > https://ift.tt/3d6eqIn > > > > Michael Habig >
via IFTTT
0 notes
Text
The Value of W, or, Interdisciplinary Engagements on Culture
OCTOBER 31, 2018
LAST SPRING, I attended a conference in New Mexico featuring evolutionary biologists working on a new research program they have been calling the “extended evolutionary synthesis” (EES). The program aims to go beyond the so-called “modern synthesis” of the mid-20th century, which joined Darwinism to Mendelian genetics, whose mathematical formulations could be simply and straightforwardly expressed. Biologists involved in the EES have been calling for a broader and less reductive view of evolution, unrestricted to Mendelian genes. In particular, they have been addressing the modern synthesis’s paucity of information about developmental biology. These EES revisionists are interested in feedbacks: in how developmental processes, along with ecological and even cultural ones, feed back into one another, into genetic and other forms of inheritance, and therefore into evolution. While the modern synthesis proposes that epigenetic, developmental, ecological, and cultural processes are all products of evolution, the EES claims they are causes as well as products.
The roots of this movement extend back to the early 1970s, to the work of Richard Lewontin at Harvard, and Luigi Luca Cavalli-Sforza and Marcus Feldman at Stanford, among others. But the reigning, reductive neo-Darwinist paradigm — in other words, the modern synthesis — remains well entrenched, and its defenders staunch in its support. Only in the last 25 years or so has the more expansive vision of the EES slowly begun — against much resistance — to establish itself in mainstream biology.
As part of this development, EES biologists have been increasingly interested in culture, among other forms of transformation and transmission, and so have welcomed the input of humanists, including philosophers and historians of science like me, whose job it is to study and understand culture. Taking part in their conversations has in turn informed my own work in the history of evolutionary theory.
Still, I experienced a moment of comical culture shock at the recent meeting I attended. A biologist wrote an equation on the whiteboard in which one of the variables was a “w.” He then circled the w, explaining that it represented “culture,” and pointed out that under certain conditions, the value of “w” would tend toward zero, while under other conditions it would tend toward 100. “Perhaps,” I thought, “we don’t mean quite the same thing by ‘culture.’” To a humanist, or anyway to this one, “culture” is an abstract noun encompassing many things of many kinds: processes, objects, habits, beliefs both explicit and implicit. It seems a category mistake to think that we can represent such a welter by a single variable, or that the whole jumble could act as a discrete thing having a single quantifiable effect on some other discrete thing. Could we say, for example, that in a given society, “culture” influences “politics” by some quantifiable amount x? Could we say that “the arts” has a y-percent effect on birth rate or life expectancy?
As it turned out, I had somewhat misunderstood the situation. When I expressed a certain dubiousness about representing “culture” with a single variable, an EES biologist explained to me that the variables standing for “culture” in biologists’ mathematical models are not meant to denote the entire Gestalt, but rather quantifiable bits of culture: a single behavior, for example, that might be taught, learned, transmitted, or counted, and whose effects on survival and reproduction can be measured and modeled. Perhaps these individual culture variables might in principle add up to a single, overarching W, but for the moment, no one claims to be able to make that summation. For now, we can simply use the little w’s to build discrete cultural bits or forms into an evolutionary model. This seems to me more credible, but it still assumes that we can meaningfully represent cultural forms as quantifiable bits, and that this will add more to our understanding of the role of cultural forms in evolutionary processes than simply trying to describe this role in qualitative terms. I can’t help wondering if that’s a sound assumption.
Of course I’m by no means the first to raise the question, nor indeed have such objections been confined to humanists. Lewontin himself, together with the historian Joseph Fracchia, argued in a 1999 paper against the idea of cultural evolution. They wondered whether conceptualizing entities like “the idea of monotheism” as “cultural units” begged crucial questions — for example, how can we count up these units in a population, and what are their laws of inheritance and variation? Fracchia and Lewontin maintained that there could be no such general laws because cultural phenomena, unlike atoms and molecules, differ from one another in their properties and dynamics of transmission and change. “There is no one transhistorical law or generality,” they contended, “that can explain the dynamics of all historical change.” [1] Marcus Feldman disagreed, albeit not specifically with regard to the existence of general laws explaining the dynamics of all historical change; rather, he defended the notion of “observable units of culture,” which he did not associate with grand organizing ideas such as monotheism. An example of an observable cultural unit for Feldman is a behavior or custom that follows statistical rules of transmission, and that can therefore be a legitimate object of mathematical study. [2]
The biologist with the “w” variable and I were thus reenacting an intellectual confrontation that has been going on for decades. As is often the case in longstanding debates, we actually agree on the essentials: that nature and culture are at bottom made of the same stuff — in fact, of one another — which no humanistic or scientific inquiry can legitimately disregard. Evolutionary theory must encompass cultural processes just as human history must encompass biological ones. But, despite our deep accord, this biologist and I are thinking incommensurately about methods, about how to put our two fields into communication. His method is mathematical modeling, and mine is thick description. These are diametrically opposite in trajectory, one abstractive and reductive, the other concretizing and expansive. While I understand and admire these biologists’ conclusions, I keep wondering: Why these methods? Why mathematical modeling? By which I mean, what function do biologists intend their mathematical models to serve? Are they meant to prove claims about evolution? Or rather to express, represent, or advocate certain interpretive views of evolutionary processes? If the latter, why choose this particular means of expression, representation, advocacy? These will surely seem naïve questions to any biologist reading this. But I have learned from teaching college freshman and sophomores that naïve questions from untrained newcomers can be the hardest and most useful, which emboldens me to ask mine.
¤
Kevin Laland, author of Darwin’s Unfinished Symphony, was an organizer of the conference I attended and is a leader of the Extended Evolutionary Synthesis research program. Reading his important and heartfelt book, the to-date summary of a groundbreaking career, I had similar feelings as I did at the conference. Uppermost among these is heated agreement: Laland’s essential tenets seem to me profoundly right, some indeed incontrovertible. These include the precept that cultural practices — in particular teaching, imitating, and copying — are causes as well as results of evolution; that in mammals and especially humans, such cultural practices have accelerated evolutionary development by constantly creating “new selection regimes” in a process that Laland, citing evolutionary biologist Allan Wilson, calls “cultural drive”; and that, accordingly, in humans especially, there has been a “gene-culture coevolutionary dynamic.” The first of these — that cultural practices are causes as well as results of evolution — seems to me incontrovertible, but more like a first principle than like an empirical result. Cultural practices must be causes as well as results of evolution because any result of the evolutionary process becomes a feature of the world of causes shaping the continuation of that process.
The other principal tenets — such as “cultural drive” and “gene-culture co-evolution” — are not quite first principles, but they seem to me ways of understanding how the feedback-loop of evolution encompasses cultural forms. To express these ways of understanding in the language of mathematical modeling seems fine, if one likes to do that, but no more definitive than expressing them in words. This is because a mathematical model, like a verbal description, contains many layers of interpretation. This is not a criticism: interpretation is essential to (and ineradicable from) any attempt to understand the world. But insofar as a mathematical model is taken to prove rather than to argue or represent, that’s where I think it can mislead.
Laland has devoted his career to pioneering work against reductive, simplistic, and dogmatic accounts of evolution, building brick by brick a sound case for the richer and more complex vision of the EES. Darwin’s Unfinished Symphony is a record of his resounding success. But while he has been constructing this revisionist scientific theory, he has often supported it by traditional methods. An example is his game-theoretic tournament to study social learning. After offering several examples of social learning in animals — such as Japanese macaques who learn from one innovative macaque to wash their sweet potatoes before eating them, and fish who learn from one another where the rich feeding patches are located — Laland asks what might be the best “social learning strategy.” He explains that the “traditional means to address such questions is to build mathematical models using, for instance, the methods of evolutionary game theory.”
Game theory became a standard model in evolutionary biology in the early 1970s with the work, notably, of the British theoretical evolutionary biologists W. D. Hamilton and John Maynard Smith, along with the population geneticist George R. Price. Hamilton, Price, and Maynard Smith developed a game-theoretic approach to modeling the behaviors of organisms in the struggle for survival. Their work was foundational to the neo-Darwinist, gene-centric program that Laland has devoted his career to challenging. In this gene-centric view, all higher-order entities — individual organisms, their behaviors and interactions — are epiphenomenal, controlled by and reducible to genes, so that any apparent agency or intention on the part of an organism is illusory. Organisms survive if they happen to achieve an optimal state of genetic affairs, one that maximizes some function for greater reproductive success. They die out when they fail to do so. Maynard Smith accordingly emphasized that his technical definition of “strategy” was strictly behaviorist. “Nothing,” he maintained, “is implied about intention.” A strategy was merely “a behavioural phenotype,” in other words, “a specification of what an individual will do [in a given situation].” [3] These “strategies,” therefore, involved no ascription of internal agency, but merely outward observations of behavior. Neither observed behaviors nor any other macrolevel phenomenon could play a causal role in evolution according to this school of thought.
Maynard Smith’s approach has inspired the most reductive of neo-Darwinists. For example, Richard Dawkins has adapted it to his own theory of gene functioning, emphasizing that the “strategies” in question are behaviorally defined and do not require the ascription of consciousness, let alone agency, to the strategic agent. Dawkins indeed refers to “unconscious strategists,” the deliberate oxymoron encouraging the reader to accept these apparent ascriptions of agency to genes as radical denials of any such agency. [4] Neither behaviors, nor agency, nor consciousness, nor culture operates causally at any level of Dawkins’s picture; all reduces to just gene functioning.
Game-theoretic modeling has been a hallmark of neo-Darwinist reductionism and, specifically, of the denial of any kind of evolutionary agency to the evolving organism. But in Darwin’s Unfinished Symphony, Laland describes how he and his collaborators used game theory in an innovative way, to design a virtual world in which they hosted a tournament. The game involved virtual “organisms” or “agents” engaging in a hundred “behavior patterns,” with varying rates of success resulting in greater or lesser “fitness” (i.e., survival and reproduction). The game also included three different “moves” — “innovate,” “observe,” and “exploit” — representing different phases of asocial or social learning. More than a hundred people of various ages and backgrounds took part in the game. Unlike in Maynard Smith’s applications of game theory to evolution, Laland and his collaborators were not looking for an optimum in the form of a single function or property to be maximized. They did not pre-judge what had to happen in order for an organism to win the competition. Rather, they set the competitors loose and waited to see who would triumph. The winning strategy was an unpredictable, complex mix of behaviors, although it did represent an overall optimum solution composed of behavioral bits.
Analyzing the winning strategy, Laland concludes that observing and copying are tremendously valuable, much more so than innovating on one’s own except “in extreme environments that change at extraordinarily high rates,” which must be rare in nature. The conclusion is persuasive, but the tournament seems to me more a way of expressing than of proving this point: the virtual agents and their behaviors and strategies of course constitute an interpretive representation of natural processes. They are not drawn in pastels or composed in prose, but the fact that they are programmed on a computer makes them no less a representation.
¤
To elaborate further, consider an experiment Laland describes, performed with his postdoctoral student Hannah Lewis. Laland explains that to model the effects of high-fidelity transmission of information on the longevity of cultural forms or “traits” in a population, he and Lewis “assumed that there are a fixed number of traits that could appear within a group through novel inventions and that are independent of any other traits within a culture. We called these novel inventions ‘cultural seed traits.’ Then, one of four possible events could occur”: a new seed trait could be acquired by novel invention; two traits could be combined to produce a new one; one trait could be modified; or a trait could be lost.
This model, in its relation to real cultural forms, seems to me the equivalent of a Cubist painting. Cultural “traits” that are independent of one another occur no more often in nature than young ladies with perfectly geometrical features distributed all on one side of their two-dimensional heads. Likewise for the separate and distinct occurrence of novel invention, combination, modification, or loss of cultural forms. These processes travel in the real world as aspects of a single organic entity and not as separate blocks. Of course, I’m not opposed to representing cultural forms in these Cubist terms any more than I’m opposed to Picasso’s portraits of Dora Maar. Representations should, though, declare themselves as such.
Mathematical models are interpretative from the get-go. Again, let me be clear that I think that’s fine — indeed, inevitable — because interpretation is ineradicable from any attempt to understand the world. Indeed, some scientists describe their use of mathematical models in these very terms. The theoretical physicist Murray Gell-Mann warned that we must be careful, regarding models, “not to take them too seriously but rather to use them as prostheses for the imagination, as sources of inspiration, as acknowledged metaphors. In that way I think they can be valuable.” [5] Feldman, who pointed me to Gell-Mann’s characterization of models as “prostheses of the imagination,” added that “insofar as the model assists in the interpretation, then it has value.” [6] On another occasion, Feldman told an interviewer, “[p]eople who make models for a living like I do don’t actually believe they’re describing reality. We aren’t saying that our model is more probable than another model; we’re saying it exposes what is possible.” [7]
I have no trouble believing in mathematical modeling as a powerful form of metaphor, representation of the possible, or prosthesis for the imagination. But mathematical modeling does have a distinctive feature that sets it apart from other interpretive modes: notwithstanding Gell-Mann and Feldman, it tends to disguise itself as proof rather than representation. Would it be possible for it to come right out of the positivist closet? To put my point another way, culture plays as crucial a role in evolutionary theory as it does in evolution. Culture plays as crucial a role in science as it does in nature. Wouldn’t a scientific method that unapologetically declared itself as interpretive and representational be in keeping with Laland’s revolutionary program to write cultural forms into evolutionary theory?
Mathematical modeling, like any mode of interpretive analysis, also has its limitations and pitfalls. For example, it brings a tendency I’ll call “either/or-ism”: a tendency to represent as separate and discrete, the better to count them, things that are in fact mixed and blended. Darwin’s Unfinished Symphony lists as discrete alternatives, for example, animals learning innovations socially from one another versus inventing them independently; the cultural drive hypothesis operating through natural selection on social learning proficiency versus social learning incidence; humans being more accomplished than other primates due to “chance factors” or because of a “trait or combination of traits that were uniquely possessed by our ancestors”; that high-fidelity transmission of information might have been achieved by our ancestors through language or alternatively through teaching; learning a skill such as stone-knapping to make a cutting tool by reverse-engineering from a finished sharpened flake, or else by imitation, or else by various forms of non-verbal teaching; or else by verbal teaching; and young individuals acquiring skills either asocially by trial and error, or else socially by copying, or else socially by being taught by a tutor “at some cost to the tutor.” In each of these cases, “both, and” seems more plausible to me than “either, or.” (Additionally, in the last case, must teaching involve a cost to the tutor? In my experience, teaching is often a win-win process, a non-zero-sum game, in which the teacher learns at least as much as the pupil, rather than a donation by the teacher to the pupil. Perhaps the sort of teaching that humans do is qualitatively different from the sorts that other animals do: a teacher macaque might not derive the same intellectual benefits from teaching to compensate for the loss of time that could be spent eating or reproducing. But I wonder if that’s necessarily true in all cases of nonhuman teachers.)
Yet Laland’s conclusions are extremely persuasive. Their persuasiveness overwhelms my failure to believe in a proof-value for the mathematical models. He concludes that natural selection favors those who copy others efficiently, strategically, and accurately; that nonhuman species lack cumulative cultures because of their “low-fidelity copying mechanisms”; that teaching evolves where the benefits outweigh the costs; and that language first evolved to teach close kin. I can believe in these conclusions, not as proven by the tournament-experiment, or the cultural-trait-transmission model, or the other mathematical models, but as interpretively, argumentatively presented by these models. I think this is because Laland’s conclusions are based on the kind of profound knowledge that comes only from a wealth of direct experience and — yes — keen, richly informed interpretation. Alongside the mathematical models are descriptions drawn from experiments and observations, some extending over decades.
For example, Laland describes several series of experiments designed to show that fish can learn from one another, and to investigate how and under what conditions they do so. In one set of experiments, Laland and his students and collaborators trained guppies to take certain routes to find rich food supplies, then observed other untrained guppies, in various conditions, learn from their trained fellows. In one variation, the experimenters trained the demonstrator fish to swim directly up narrow vertical tubes to reach their meal; this was a highly esoteric skill that no fish figured out on its own, without training, but the guppies did readily learn it from one another. In another series of experiments, the experimenters offered certain stickleback fish rich feeding patches and others poor ones, while observer fish watched from a distance; the humans then observed the observer fish to see whether and what they learned.
Such experiments, Laland reports, have established certain social tendencies in fish. These include “a tendency to adopt the majority behavior,” “copying the behavior of others when uncertain,” and “disproportionately attending to the behavior of groups.” Such social tendencies, once established, must surely enter into any legitimate evolutionary picture of fish. More generally, the principle that many animals are social, and that their sociality necessarily plays a role in the evolutionary process, has the retrospective obviousness of all grand, organizing ideas once stated, a most notable example being the idea of natural selection itself, whose retrospective obviousness led T. H. Huxley, upon reading the On the Origin of Species, to figuratively smack his forehead, exclaiming: “How extremely stupid of me not to have thought of that!” [8] Such grand, organizing ideas, which create conceptual sea-changes that render them retrospectively (but only retrospectively) obvious, can emerge only from richly informed interpretative analysis.
Darwin’s own method was explicitly so. He described natural history as a form of deeply interpretive historical scholarship. The geological record, he said, was a collection of fragments of the most recent volume of “a history of the world imperfectly kept, and written in a changing dialect.” He urged people to join him in considering natural history in these terms: to “regard every production of nature as one which has had a history” to be pieced together by interpretation of scant evidence. Darwin promised that this approach would be its own reward: “[W]hen we thus view each organic being, how far more interesting, I speak from experience, will the study of natural history become!” [9] Laland’s evolutionary science, as portrayed in Darwin’s Unfinished Symphony, might as well come right out and declare itself as such: it is precisely that “far more interesting” study.
¤
Jessica Riskin is a history professor at Stanford University, where she teaches courses in European intellectual and cultural history and the history of science. She is the author, most recently, of The Restless Clock: A History of the Centuries-Long Argument Over What Makes Living Things Tick (2016).
¤
[1] Joseph Fracchia and R. C. Lewontin, “Does Culture Evolve,” in History and Theory Vol. 38, No. 4, Theme Issue 38: The Return of Science: Evolutionary Ideas and History (Dec., 1999), pp. 52–78, on pp. 60, 72.
[2] Marcus W. Feldman, “Dissent with Modification: Cultural Evolution and Social Niche Construction,” in Melissa J. Brown, ed., Explaining Culture Scientifically (Seattle: University of Washington Press, 2008), Ch. 3, on p. 58.
[3] John Maynard Smith, Evolution and the Theory of Games (Cambridge: Cambridge University Press, 1982), 5, 10.
[4] Richard Dawkins, The Selfish Gene (1976), 30th anniversary ed. (Oxford: Oxford University Press, 2006), 229.
[5] Murray Gell-Mann, “Plectics,” in John Brockman, ed., Third Culture: Beyond the Scientific Revolution (New York: Touchstone, 1995), Ch. 19, on p. 324.
[6] Marcus Feldman, in conversation, August 2018.
[7] Feldman, quoted in Elizabeth Svoboda, “Finding the Actions that Alter Evolution,” in Quanta Magazine, Jaunary 5, 2017, https://www.quantamagazine.org/culture-meets-evolution-the-marcus-feldman-qa-20170105/.
[8] Thomas Henry Huxley, “On the Reception of The Origin of Species” (1887), in The Life and Letters of Charles Darwin, edited by Francis Darwin (New York: D. Appleton, 1896), 1:533–58, on p. 551.
[9] Charles Darwin, On the Origin of Species by means of natural selection, or the preservation of favoured races in the struggle for life (London: John Murray, 1859 [1st ed.]), 310–311, 485–486.

Source: https://lareviewofbooks.org/article/the-value-of-w-or-interdisciplinary-engagements-on-culture/
0 notes
Text
New Tools for Hop Cytogenomics: Identification of Tandem Repeat Families from Long-Read Sequences of Humulus lupulus
Hop (Humulus lupulus L.) is known for its use as a bittering agent in beer and has a rich history of cultivation, beginning in Europe and now spanning the globe. There are five wild varieties worldwide, which may have been introgressed with cultivated varieties. As a dioecious species, its obligate outcrossing, non-Mendelian inheritance, and genomic structural variability have confounded directed breeding efforts. Consequently, understanding genome evolution in Humulus represents a considerable challenge, requiring additional resources, including integrated genome maps. In order to facilitate cytogenetic investigations into the transmission genetics of hop, we report here the identification and characterization of 17 new and distinct tandem repeat sequence families. A tandem repeat discovery pipeline was developed using k-mer filtering and dot plot analysis of PacBio long-read sequences from the hop cultivar Apollo. We produced oligonucleotide FISH probes from conserved regions of HuluTR120 and HulTR225 and demonstrated their utility to stain meiotic chromosomes from wild hop, var. neomexicanus. The HuluTR225 FISH probe hybridized to several loci per nucleus and exhibited irregular, non-Mendelian transmission in male meiocytes of wild hop. Collectively, these tandem repeat sequence families not only represent unique and valuable new cytogenetic reagents but also have the capacity to inform genome assembly efforts and support comparative genomic analyses. http://dlvr.it/RPLjtB
0 notes
Last Seen Blogs

feliciaelizabeth94
Felicia Elizabeth

theauragirl
the aura girl

possiblyfreddy
18|CA|🎼


yoursidehismain
Untitled