#massopoda
Text

opposite to conventional representation, massopoda sculpture of the late Triassic period was often brightly painted. Imagine the ancient city roads flanked with kneeling Kholumolumo Ellenbergerorum warriors such as this one
#art#painting#digital art#digital painting#anthro#furry#sculpture#massopoda#kholumolumo ellenbergerorum#dinosaur#saurian#sauropod#scalie#anthro art#furry art#fantasy#statue#warrior#dynasoar5#ssohardd
1K notes
·
View notes
Text
It Came From The Trash Heap (We Don't Talk About Kholumolumo)
A wastebasket taxon is what happens when species can't be easily classified and instead get hurled into a "catch-all" category.
…But that's not the only kind of taxonomic tangle that can befall a new discovery.
When a scientific name is assigned to a new species, but it isn't given a corresponding formal description and type specimen, it becomes a nomen nudum – a "naked name". Without a proper description and assigned holotype the name isn't valid, and the new species isn't technically accepted by the wider scientific community.
This has even happened to some surprisingly famous names. In the 1920s Velociraptor mongoliensis was briefly given the nomen nudum "Ovoraptor djadochtari" before getting its much more familiar name when it was officially described. Meanwhile the giant pterosaur Quetzalcoatlus northropi was stuck as a nomen nudum for decades, only finally getting a proper published description in 2021.
And there's another particular long-standing nomen nudum that became mildly infamous – "Thotobolosaurus", the "trash heap lizard".

Discovered next to a literal trash pile in the village of Maphutseng in Lesotho, a few scattered and broken bones of this "prosauropod" sauropodomorph dinosaur were first found in 1930. But it wasn't until the mid-1950s that a more extensive bonebed began to be unearthed at the site, and over the next decade over 1000 fossil fragments were collected.
In the mid-1960s the remains were initially classified as belonging to Euskelosaurus browni (which is now considered to be a wastebasket taxon), but just a few years later in 1970 the "Maphutseng Beast" was re-evaluated as a species new to science. It was referred to as "Thotobolosaurus mabeatae" – based on the local name of the discovery site, "Thotobolo ea ‘Ma-Beata" (trash heap of Beata’s mother) – but this name was never actually formally published.
Despite "Thotobolosaurus" being an undescribed nomen nudum it nonetheless went on to be repeatedly referenced in scientific literature over the next few decades, and appeared in several popular dinosaur books (even as recently as 2020!).
In the mid-1990s it was alternatively named "Kholumolumosaurus ellenbergerorum" in a Ph.D. dissertation, with this name derived from the kholumolumo, a reptilian creature in Sotho mythology, and the Ellenberger brothers who worked on the site. But this also didn't count as a formal publication and instead became a second nomen nudum for the species.
Eventually, 90 years after the first bones were found and 50 years after the debut of the name "Thotobolosaurus", this long-neglected sauropodomorph was finally given a proper published full anatomical description in 2020.
And it also got a third name, this time officially valid, based on the second one from the 1990s: Kholumolumo ellenbergerorum.
For something associated with trash for so long, Kholumolumo is actually now one of the most completely-known prosauropods. At least five different individuals were present in the collected fossil material, possibly as many as ten, and between them most of the full skeleton is represented – with the exception of the skulls, which are only known from a couple of small fragments.
We now know Kholumolumo was rather heavily-built, with chunky limb bones and unusually short shinbones. It would have been one of the biggest animals around in the Late Triassic (~210 million years ago), measuring at least 9m long (~30') and weighing around 1.7 tonnes (1.9 US tons), but despite its size it seems to have still been bipedal.
Due to the highly disarticulated nature of the bones the fossil site may have been a "bone accumulation area", a place where dismembered bits and pieces of different carcasses were regularly carried to be eaten by a predator or scavenger – essentially a trash heap, fittingly enough. A couple of "rauisuchian" teeth have actually been found among the remains, which might indicate what was chomping on these particular Kholumolumo.
———
Nix Illustration | Tumblr | Twitter | Patreon
#it came from the wastebasket#nomen nudum#taxonomy#kholumolumo#thotobolosaurus#prosauropod#massopoda#sauropodomorpha#dinosaur#paleontology#art#science illustration#paleoart#palaeoblr
290 notes
·
View notes
Text

Day 5: Massospondylus carinatus.
Based on skeletal design by Scott Hartman.
#massospondylus#massospondylidae#massopoda#plateosauria#sauropodomorpha#saurischia#dinosauria#jurassic june#early jurassic#paleoart#coloredpencil#my art
11 notes
·
View notes
Text
Melanorosaurus readi

By Tas Dixon
Etymology: Black Mountain Reptile
First Described By: Haughton, 1924
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Melanorosauridae
Time and Place: From 210 until 201 million years ago, possibly into the Jurassic, from the Norian through the Rhaetian (and possibly into the Hettangian) of the Late Triassic

Melanorosaurus is known from the Lower Elliot (and, maybe, the Upper Elliot) Formation of South Africa
Physical Description: Melanorosaurus was a later Sauropodomorph, the group of dinosaurs that includes Sauropods and their close relatives (aka, “Prosauropods”). Melanorosaurus was technically not a Sauropod, but it does highlight how these quadrupedal behemoths evolved from basic dinosaur precursors. It was quadrupedal, but with awkward front feet still retaining hand-like qualities of its forefathers. The hindlimbs were more sturdy, and in general the pillar-like proportions of the limbs helped to hold up its great weight. It was probably up to 8 meters long and 2 meters tall, making it one of the heaviest animals in its environment - weighing about 1.3 tons. It had a pointed, triangular snout, and teeth like earlier Prosauropods rather than proper Sauropods. It had a short neck and long tail, with a very thick trunk. As for external appearance, there is a question - it was small enough, just, to still have fluff, but it also was big enough to have lost it in the interest of keeping cool. Given it lived in a particularly hot climate, it makes sense that some - if not all - of the warm fluff of its ancestors may have been shed off. That said, it also lived southward - so it’s possible that the Elliot had a cooler environment than other places of the Late Triassic. All in all, the fluff status of Melanorosaurus is a question, so here we present it fluffy to some extent, since most interpretations of it are scaly.
Diet: Melanorosaurus would have been an herbivore, but it may have fed occasionally on small animals to supplement its diet, especially since Melanorosaurus had the teeth of its omnivorous precursors.

By Ripley Cook
Behavior: Melanorosaurus was a smaller Sauropodomorph, so it probably would have taken care of its young (like its close relative Massospondylus) in nests and potential family structures. These nests are hypothetical, however, so we can’t say what sort of lifestyle Melanorosaurus would have lead in this regard. It probably would have stuck together in herds for safety from large contemporaneous predators, and moved together across the dry landscape of their environment looking for new sources of food. Given they managed to survive the end-Triassic extinction (probably), this was clearly a successful strategy to some extent. Their short necks means that they probably would have had to feed mainly on low-lying vegetation, though it is entirely likely that they could have reared on their hind limbs in order to reach higher sources of food. Since they still had some traits of bipedality (likely vestigial), this seems more likely than not.
Ecosystem: The Elliot Formation was a highly arid sub-tropical desert, filled with hearty conifers (which did a lot of the desert-plant jobs before cacti evolved) lining seasonal rivers that would dry up come the long harsh season. It was cooler than other places in the Triassic, but still quite hot and harsh, making it a baking environment for the creatures that lived there. The exact composition of each environment - the Lower Triassic part of the environment and the Upper Jurassic part - is difficult to determine, because the levels tend to be hard to define, but some work has been done on this in recent years. Other Sauropodomorphs included Plateosauravus, Eucnemesaurus, and Bikanasaurus - so, some not very well preserved species, making Melanorosaurus an exception in this region. There was also the Mammaliaform Elliotherium and the Dicynodont Pentasaurus. However, literally every other example of creature seems to be from the Upper Elliot, so it is uncertain what the predators of Melanorosaurus would have been.

By José Carlos Cortés
Other: Melanorosaurus is such an almost-Sauropod that its description and study often threatens to redefine exactly what it means to be a Sauropod or… not. As such, Melanorosaurus is literally defined out of Sauropoda, with Sauropods defined as those members of the sauropod-y group more closely related to Saltasaurus than to Melanorosaurus. That said, Melanorosaurus has a significant amount of similarities to later Sauropods, one of the weirder and more magnificent experiments of Triassic dinosaurs - and a sign of the scale of dinosaurs to come after this period of experimentation ends.
~ By Meig Dickson
Sources Under the Cut
Fabrègues, P. C. d., R. Allain. 2016. New material and revision of Melanorosaurus thabanensis, a basal sauropodomorph from the Upper Triassic of Lesotho. PeerJ 4: e1639.
Galton, P. M., P. Upchurch. “Prosauropoda”. In D. B. Weishampel, P. Dodson, & H. Osmólska (eds.). 2004. The Dinosauria (second edition). University of California Press, Berkeley 232 - 258.
Galton, P. M., J. Van Heerden, A. M. Yates. 2005. Postcranial Anatomy of Referred Specimens of the Sauropodomorph Dinosaur Melanorosaurus from the Upper Triassic of South Africa. Tidwell, V. & K. Carpenter (eds.)Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press.
Gauffre, F. X. 1993. The most recent Melanorosauridae (Saurischia, Prosauropoda), Lower Jurassic of Lesotho, with remarks on the prosauropod phylogeny. Neues Jahrbuch für Geologie und Paläontologie Monatshefte. 1993 (11): 648 - 654.
Haughton, S. H. 1924. The fauna and stratigraphy of the Stormberg Series. Annals of the South African Museum 12: 323 - 497.
McPhee, B. W., J. N. Choiniere, A. M. Yates and P. A. Viglietti. 2015. A second species of Eucnemesaurus Van Hoepen, 1920 (Dinosauria, Sauropodomorpha): new information on the diversity and evolution of the sauropodomorph fauna of South Africa's lower Elliot Formation (latest Triassic). Journal of Vertebrate Paleontology 35(5):e980504:1-24.
McPhee, B. W., E. M. Bordy, L. Sciscio, J. N. Choiniere. 2017. The sauropodomorph biostratigraphy of the Elliot Formation of southern Africa: Tracking the evolution of Sauropodomorpha across the Triassic–Jurassic boundary. Acta Palaeontologica Polonica 62 (3): 441 - 465.
Paul, G. S. 2010. The Princeton Field Guide to Dinosaurs. Princeton University Press: 170.
Yates, A. M. 2007. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodmorpha: Anchisauria), in Barrett, P. M. & D. J. Batten. Special Papers in Paleontology 77: 9 - 55.
Yates, A. M. 2010. A revision of the problematic sauropodomorph dinosaurs from Manchester, Connecticut and the status of Anchisaurus Marsh. Palaeontology 53 (4): 739 - 752.
#melanorosaurus#melanorosaurus readi#palaeoblr#triassic#dinosaur#prosauropod#sauropodomorph#reptile#prehistoric life#paleontology#triassic madness#triassic march madness
337 notes
·
View notes
Note
Do you have any resources that could give me a good overall explanation of dinosaur classification? I’m pretty new to paleoblr and I get mixed up with all of the names floating around. Thanks and sorry if I bothered you!
I don’t - but I’ll be happy to write a bit!
1. What is a “Dinosaur”?
This may seem obvious, but it’s a bit more subtle. To scientists, “Dinosauria” is a group containing the most recent creature that was the ancestor of T. rex, Triceratops, and Brontosaurus, and everything descended from that ancestor. This means that birds, as descendants of dinosaurs, are included under that definition, while pterosaurs and most mesozoic sea reptiles like plesiosaurs are not.
This kind of group is called a “node-based group”, and is written as (Tyrannosaurus+Triceratops+Brontosaurus).
There’s also “stem-based groups”, which is “everything closer related to X than to Y”. A good example of this is reptilia, which is something like (Alligator>Homo) [that is, everything closer to alligators than to people!].
2. 3 main groups
There’s three main groups of dinosaurs - theropods, sauropodomorphs, and ornithischians. The simplest (though not always accurate) way of thinking about these is that theropods are two-legged meat-eaters, sauropodomorphs are long-necked plant eaters, and ornithischians are beaked plant-eaters.
For a long time it was accepted that theropods and sauropodomorphs were each others’ closest relatives, in a group called saurischia, and that this group was in turn the closest relatives of ornithischians. Recent analyses show that this may not be entirely accurate - it may be that theropods and ornithischians are united in a group called ornithoscelida, and that sauropodomorphs are the closest relative of this group. There’s good reasons to think each is true, and there’s going to need to be more research done in the future, and hopefully more fossils will straighten things out.
3. Ornithischia
Nearly all ornithischians have three things in common:
They’re mainly herbivores
They have a special bone on their lower jaw called a predentary that formed part of a beak
Part of their hips faces backwards, allowing more room for guts (important because plants are hard to digest!)
There’s three main groups of ornithischians, as well as a bunch that don’t really fit into any of those groups.
The most significant of these “oddballs” are the heterodontosaurs, a group of early ornithischians that mainly lived in the jurassic and triassic periods. They’re generally small (60-200cm in length) two-legged omnivores or herbivores that had big fangs that were probably used for display. They’re kind of the weird cousins of all other ornithischians.
Thyreophorans
This literally means “shield bearers”, and as you might expect it includes the armoured dinosaurs like Stegosaurus and Ankylosaurus. It also includes some weird early forms like Scutellosaurus. It’s defined as (Ankylosaurus>Triceratops).
>Eurypods
This is specifically (Stegosaurus+Ankylosaurus), so it’s contained within thyreophora.
>>Ankylosauria
This is (Ankylosaurus>Stegosaurus), and contains the most heavily armoured dinosaurs. It’s divided into Ankylosauridae (ones with tail clubs), Nodosauridae (which have bigger shoulder spikes), and possibly also Polacanthidae (which have more sticky-up spikes, but also might just be nodosaurids)
>>Stegosauridae
This is (Stegosaurus>Ankylosaurus), and contains the familiar plated dinosaurs. It includes Stegosauridae (the big ones like Stegosaurus) and Huayangosauridae (some smaller Chinese forms).
Neornithischia
This group is defined as (Parasaurolophus>Stegosaurus/Ankylosaurus). It contains two major groups - the Marginocephalians and the Ornithopods, but also a bunch of important basal members, like Thescelosauridae, Kulindadromeus and Hypsilophodon - animals that were once thought to be ornithopods but probably aren’t.
Marginocephalia
This is (Pachycephalosaurus+Triceratops). The name means “rimmed head”, because….both major groups have big stuff around their heads.
>Pachycephalosauria
These are the “bone-headed” dinosaurs like Pachycephalosaurus. It’s (Pachycephalosaurus>Triceratops)
>Ceratopsia
This is (Triceratops>Pachycephalosaurus), and contains the beakiest of all dinosaurs. Chaoyangosaurids are frill-less, hornless, 2-legged forms; Bagaceratopsids, Leptoceratopsids, and Protoceratopsids are hornless but increasingly frilled and 4-legged groups.
>>Ceratopsidae
This is (Centrosaurus+Triceratops), and contains the big, 4-legged, horned guys. Centrosaurines usually have smaller frills with big horns around them, smaller brow horns, and bigger nose horns, while Chasmosaurines usually have bigger frills, bigger brow horns, and smaller nose horns.
Ornithopoda
This is (Parasaurolophus>Triceratops). It used to contain a bunch more stuff, but now it mostly contains just Iguanodonts, so for most purposes those are the same thing (except for some southern forms called Elasmaria that don’t come up much). Doesn’t matter as much as it used to; them’s the breaks. It includes Rhabdodontids, a weirdogroup of small bipedal guys from Europe, and Dryomorpha.
>>Dryomorpha
This is (Dryosaurus+Iguanodon). It contains Dryosauridae, a group of fast-running ornithopods, and Ankylopollexians, the group that had the famous “thumb-spikes).
>>>Styracosterna
Except for a few species, this is about the same as Ankylopollexia. It contains a bunch of species, most of which used to just be called Iguanodon, as well as the Hadrosauriformes, which contains the Hadrosauroids, which contains the Hadrosauromorphs, which contains the Hadrosaurids (whew!)
>>>>Harosauridae
These are the “Duck-bills”. It contains the Lambeosaurines, which had big long hollow crests they could use to HONK !, as well as the Hadrosaurines, which didn’t have hollow crests.
4. Sauropodomorpha
This group is mainly made up of long-necked plant eaters. It starts off with a bunch of things we used to call “Prosauropods”, but now call…….basal sauropodomorphs. It may include Herrersauridae (Pedants be quiet), a group of early, early predators. It probably includes Guaibasaurids, a group of small omnivores from the triassic. It also includes Plateosaurids, a group of larger (but still bipedal), long-necked herbivores. From here we go into Massopoda, a group that includes Massospondylids and Riojasaurids, which…are similar to plateosaurids, as well as Sauropodiformes.
Sauropodiformes is where we start to get more sauropod-y, though we still have to zoom through Anchisauria to get to actual sauropods. We’re there now.
Sauropoda
These guys are actually quadrupedal! Here we’ve got….more sliding groups. There’s a bunch of early sauropods that are quite cool, but I’m not gonna name them. The fun group is Gravisauria. This includes some early guys calls Vulcanodonts, and Eusauropods,
Eusauropods include – you guessed it! More grades. There’s some interesting features here though - Mamenchisaurids are a bunch of Chinese forms with super long necks, and I can’t say I know anything remarkable about Turiasaurs. You’ll have to talk to John about that one.
Neosauropoda
Here’s where you’ll start recognising things if you haven’t already. This is (Saltasaurus+Diplodocus), and contains the most famous sauropods. It’s split into two main groups.
>Diplodocoidea
There are the “whip-tails” (again, pedants be quiet!). It contains Dicraeosaurids, a few weird, short-necked, double sailed things, Rebbachisaurids, a group of wide-mouthed weirdos, and Diplodocids, the famous great swan-necked ones that are some of the largest dinosaurs ever.
>Macronaria
This group contains some basal forms like Camarasaurus, and two main groups (Maybe?? This is kind of a contentious area). Brachiosaurids include the giant, super-tall ones, and some little ones also. Then there’s the monster that is Somphospondyli. This contains….more grades, the Euhelopodids, and the Titanosaurs.
>>Titanosauria.
This is a real monster, let me tell you. It includes more grades – yay! – and Lithostrotia.
>>>Lithostrotia
This is where titanosaurs start getting armoured (except it’s not really that simple, since others have armour, and it may have evolved multiple times, and…..lots of stuff). It also includes the real giants like Lognkosaurs, Aeolosaurs, and Saltasaurs.
5. Theropods
These are the two-legged meat-eaters – although many are omnivorous or herbivorous! They include some early forms and Neotheropods. (From here on, except when notes, groups in big font include the rest of the groups listed below).
Neotheropods
These include the early, long-necked Coelophysoids and Dilophosaurids (which may well be more advanced possibly even Tetenurans!). This group also contains the:
Averostrans
Literally “bird beaks”, although they didn’t all have beaks. It includes the Ceratosaurs, a group that contains some weird forms, and the Abelisaurs (large, short-armed, and bulldog faced) and the Noasaurs (Small, longer arms, need a good orthodontist).
Averostrans also include the:
Tetenurans
Named after their stiffened tails, around here is where theropods lost their fourth finger. After some basal forms it includes the:
>Megalosauroids
These consist of two main groups - the heavily built Megalosaurids and the fish-specialist Spinosaurids.
Tetenurae also includes the:
Avetheropods
This consists of the Carnosaurs and the Coelurosaurs.
>Carnosaurs
These are your big sauropod killers. They include Metriacanthosaurids and Allosaurids, as well as Carcharodontosaurians. This last group is divided into Carcharodontosaurids, which includes some of the largest predators ever to walk the Earth, and the Neovenatorids, a smaller group that MAY contain the Megaraptorans.
The other group of avetheropods is the:
Coelurosaurs
This is the earliest that we can definitively say, with pretty good certainty, that all members had feathers. It includes some basal forms and the:
Tyannoraptors
….which hands-down win the coolest name competition. This group includes the Tyrannosauroids, which I’m sure need no introduction.
It may also include the famously small Compsognathids, but those may also be outside tyrannoraptora.
It also contains the:
Maniraptoriformes
This group is the earliest we can say that all members had wings. It includes the famous “ostrich dinosaurs” or Ornithomimosaurs, and the:
Maniraptora
This group is where we first see the wing-folding ability like in modern birds. It includes the tiny, 1-fingered Alvarezsauroids and, at the other end of the spectrum, the giant, long-necked, pot-bellied, wickedly-clawed, plant-eating Therizinosaurs. It also includes the:
Pennaraptors
This group is the earliest place we see true vaned feathers. It includes the Oviraptorosauria, a group of typically beaked and crested omnivores and herbivores, as well as the
Eumaniraptors
AKA Paravians - it’s the difference between a stem-based and node-based group, but they’re essentially the same.
This group includes the really birdy things, like Anchiornis and the dragon-like Scansoriopterygids (really rolls off the tongue after some practice, I promise!). It also includes the famous “Raptors” – the Dromaeosaurids. There’s also the sickle-clawed but more omnivorous or herbivorous Troodontids, famous for their brains. This latter group may be a sister group to Dromaeosauridae, or it may be closer to:
Avialae
This is (Passer>Troodon, Deinonychus). It’s what most scientists would call “birds”. It includes some early forms exemplified by Archaeopteryx and Jeholornis, as well as:
Pygostylia
This is birds with shortened, fused tails. It’s (Confuciusornis+Passer), and includes the cool streamer-tailed Confuciusornithids. It also includes:
Ornithothoraces
This group of birds is (Enantiornis+Passer). It includes the very successful and widespead (but now extinct) bird group called the Enantiornithines. It also includes the
Euornithines
This is where we first see birds with modern-style tails. It includes some basal forms at the
Ornithuromorphs
This group contains some early groups, the Hongshanornithids and the Songlingornithids, and the:
Ornithurans
This is sort of the “last burst” before we get to true birds! It includes the seagull-like Ichthyornis and the seal-like Hesperornithines. From here on out, everything is included in the:
Neornithines
We’ve made it! This is true birds - no teeth here. From here we’re divided into – What, did you think we were done? – we’re divided into the Palaeognaths and the Neognaths.
Palaeognaths include giant flightless birds like ostriches and emus, as well as the kiwi and the chicken-like Tinamids.
Neognaths
This contains all the familiar birds.
One major group is the Galloanserans.
These include the Odontoanseres, possibly* including the albatross-like Pelagornithids as well as the famous “horse-eating” (but actually vegetarian) Gastornithids and Dromornithids, and ducks, geese, swans, and screamer birds in Anseriformes.
*Pelagornithids may be more basal galloanserans or neognaths
Galloanserae also includes Pangalliformes, which consists of megapode fowl, chickens, turkeys, pheasants, peacocks, the whole shebang.
Neognathae also includes the:
Neoaves
From here taxonomy does get a bit muddled for a while. I’ll present the two major hypotheses.
1. Columbea/Passerea hypothesis
The Columbimorphs consists of Columbiformes or pigeons and doves, Pteroclidoformes or sandgrouse, and Mesitornithiformes or Mesites.
Columbimorphs may be closest to Mirandornithes, which consists of Phoenicopteriformes or flamingoes, and Podicepidiformes or grebes. If so, this clade is called Columbea.
The rest of the birds in this hypothesis belong in a clade called Passerea.
Otidomorphs are a group consisting of Otidids or bustards, Cuculiformes or cuckoos, and Musopagoformes or turacos.
Otidomorphs may be closest to Strisores, which include Apodiformes (Hummingbirds and swifts) and Caprimulgiformes (Nightjars, owlet-nightjars, and frogmouths). In this case their clade is Otidae, not to be confused with Otididae.
The rest of the birds in this hypothesis clade together in a clade called Litoritelluraves.
Gruae may be a group consisting of Opisthocomids (Hoatzins), Charadriiformes (Gulls, terns, puffins, plovers, sandpipers), and Gruiformes (Cranes, rails).
The rest of the birds in this hypothesis clade together in an unnamed clade.
This group contains the Telluraves (more on them later!) and the:
Ardeae
This consists of Eurypygimorphs, consisting of Eurypigids (Sunbitterns) and Phaethoniformes (Tropicbirds).
Ardeae may also include:
Aequornithes
This group of waterbirds includes Gaviiformes (divers/loons), Austrodyptornithes (a clade that includes Sphenisciformes [Penguins] and Procellariiformes [Albatrosses, petrels]), Ciconiiformes (Storks), Suliformes (Boobies, gannets, cormorants), and Pelicaniformes (Pelicans, herons, ibises).
2. Columbaves Hypothesis
Strisores may be the earliest to branch off of Neoaves.
Columbimorphs may alternatively be closest to Otidomorphs, If so, this clade is called Columbaves.
The rest of the birds in this hypothesis clade together in an unnamed clade.
Gruiformes may have been the earliest to branch off in this clade.
The rest of the birds in this hypothesis clade together in an unnamed clade.
The Aequorlitornithes (Not to be confused with Aequornithes) may consist of Mirandornithes clading with Charadriiformes as a sister group to a clade between Eurypygimorphs and Aequornithes.
Also included in this unnamed clade is the
Inopaves
This may consist of Opisthocomids and Telluraves.
Telluraves – Back to your regularly scheduled program
There’s actually, to my knowledge, some degree of concensus here. It’s divided into two main groups - Afroaves and Australaves.
Australaves consists of Cariamiformes (seriemas and terror birds) and Eufalconimorphae.
>Eufalconimorphae consists of Falconiformes (Falcons, kestrels, and caracaras), as well as Psittacopasserans.
>>Psittacopasserans consist of Psittacoformes (Parrots and their ilk) and Passeriformes (Perching birds, which I’m not going into more detail on because I’d be here for a month).
Afroaves
This clade consists of Accipterimophs (New-world vultures, hawks, eagles, old-world vultures, kites), and an unnamed clade.
Unnamed clade
This clade consists of Strigiformes (owls and kin) and Coraciimorphs.
Coraciimorphs
This clade consists of Coliiformes (mousebirds), the cuckoo-roller, and Cavitaves.
Cavitaves
This clade consists of the Trogonids (Trogons and Quetzals) and the Picocoraciae.
Picocoraciae
This clade consists of Bucerotiformes (Hornbills, hoopoes, woodhoopoes) and the Picodynastornithes.
Picodynastornithes
Last one! This consists of Coraciiformes (Kingfishers, rollers, motmots, and bee eaters), and the Piciformes, or toucans, barbets, and woodpeckers.
230 notes
·
View notes
Text
Europatitan eastwoodi

By Stolp
Etymology: European titan
First Described By: Fernández-Baldor et al., 2017
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Macronaria, Titanosauriformes, Somphospondyli
Status: Extinct
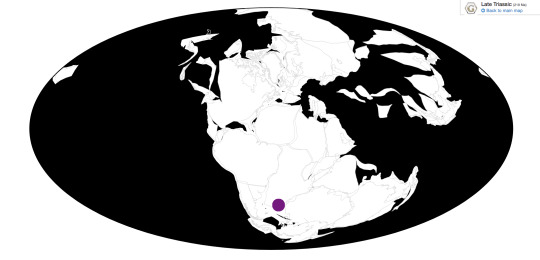
Time and Place: Around 125 million years ago, at the boundary between the Barremian and Aptian ages of the Early Cretaceous

Europatitan is known only from the Castrillo de la Reina Formation in Spain.

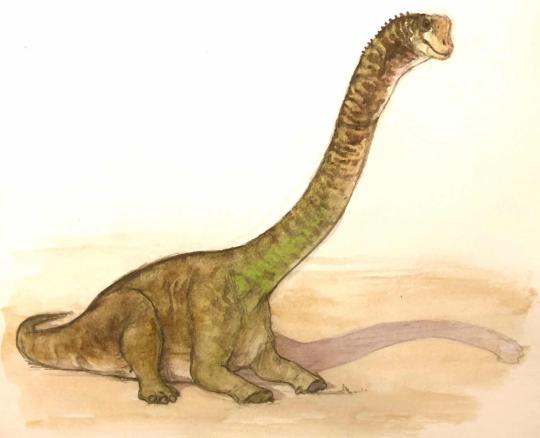
Physical Description: Europatitan would have looked a lot like other basal titanosauriformes such as Brachiosaurus. The neck vertebrae of Europatitan are especially elongate and would have been heavily pneumatized by air sacs. Although not all of the neck is known, these indicate it would have been extremely long, similar to Giraffatitan or Sauroposeidon. The body of Europatitan would have been quite wide, as its ribs indicate, and its tail fairly short. The only limb elements of Europatitan known are a couple metacarpals, but even these are enough to show it had robust forelimbs. The head of Europatitan is unknown except for a tooth, but it would have likely been similar to that of Giraffatitan.

By Scott Reid
Diet: As a sauropod, Europatitan would have been herbivorous. Its teeth were spoon-shaped, unlike the pencil-shaped teeth of diplodocoids and titanosaurs, indicating it probably had a different diet than them (which would have been useful to avoid competition with the contemporary Demandasaurus). Based on neck size, their target diet would have likely been at the tallest parts of trees.
Behavior: Europatitan would have spent much of its time eating leaves at the top of the forest tree-line as far as its neck could reach, and then moving to the next available spot for foliage. Like many sauropods, Europatitan likely spent time with others of its genera in order to reduce predation on the more vulnerable members of the group (such as sick, older, or younger individuals). While we don’t have any direct evidence for whether or not Europatitan actively cared for its young or not, the basal behavior shared among all extant archosaurs (crocodiles and birds) is some degree of parental care.

By José Carlos Cortés
Ecosystem: Europatitan’s fossils were found in the Castrillo de la Reina Formation, which in the Cretaceous would have been a floodplain, with a river flowing in from the southwest. The Castrillo de la Reina environment was also home to the rebbachisaurid Demandasaurus, the earliest rhabdodont, other indeterminate ornithopods, theropods, and the lizard Arcanosaurus. We don’t know how many of these directly lived alongside Europatitan, though, as the specific site that Europatitan was found at has only yielded theropod teeth and fragments of an iguanodont.
By Ripley Cook
Other: Fun fact: the type species of Europatitan, E. eastwoodi, is named for actor Clint Eastwood. The Good, the Bad, and the Ugly, which he starred in, was filmed near where they found the fossil.
~ By Henry Thomas and Blaire Fei
Sources Under the Cut
Dieudonné, Paul-Emile; Tortosa, Thierry; Torcida Fernández-Baldor, Fidel; Canudo, José Ignacio; Díaz-Martínez, Ignacio (2016-06-22). Farke, Andrew A. (ed.). "An Unexpected Early Rhabdodontid from Europe (Lower Cretaceous of Salas de los Infantes, Burgos Province, Spain) and a Re-Examination of Basal Iguanodontian Relationships". PLOS ONE. 11 (6): e0156251.
Fernández-Baldor, Fidel Torcida; Canudo, José Ignacio; Huerta, Pedro; Montero, Diego; Suberbiola, Xabier Pereda; Salgado, Leonardo (September 2011). "Demandasaurus darwini , a New Rebbachisaurid Sauropod from the Early Cretaceous of the Iberian Peninsula". Acta Palaeontologica Polonica. 56 (3): 535–552.
Fernández-Baldor, F.T.; Canudo, J.I.; Huerta, P.; Moreno-Azanza, M.; Montero, D. (2017). "Europatitan eastwoodi, a new sauropod from the lower Cretaceous of Iberia in the initial radiation of somphospondylans in Laurasia". PeerJ. 5: e3409.
Houssaye, Alexandra; Rage, Jean-Claude; Torcida Fernández-Baldor, Fidel; Huerta, Pedro; Bardet, Nathalie; Pereda Suberbiola, Xabier (April 2013). "A new varanoid squamate from the Early Cretaceous (Barremian–Aptian) of Burgos, Spain". Cretaceous Research. 41: 127–135.
#Europatitan#Europatitan eastwoodi#Dinosaur#Sauropod#Palaeoblr#Dinosaurs#Prehistoric Life#Paleontology#Eurasia#Prehistory#Somphospondylian#cretaceous#Herbivore#Terrestrial Tuesday
161 notes
·
View notes
Text
Barosaurus lentus

By Ripley Cook
Etymology: Heavy Reptile
First Described By: Marsh, 1890
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Diplodocoidea, Diplodocimorpha, Flagellicaudata, Diplodocidae, Diplodocinae
Status: Extinct
Time and Place: Between 150 and 149 million years ago, in the Tithonian of the Late Jurassic

Barosaurus is known from the Brush Basin Member of the Morrison Formation in South Dakota, Utah, and Wyoming. Potential specimens of Barosaurus are known from other locations of the Morrison Formation; the entire range of this habitat at the time of Barosaurus is shown below in green (with the range of Barosaurus inside of it, in blue).

Physical Description: Barosaurus in a lot of ways is a fairly typical Diplodocid sauropod - long, large, and with a distinctive whip-tail. But, when you dig under the surface, Barosaurus has nothing truly “typical” about it. The neck of Barosaurus is next-level in its length - and the tail is ridiculous to match. In fact, the estimates of the length of Barosaurus get huge - it was probably more than twenty-six meters long, and some of the most upper estimates of Barosaurus have it at fifty meters long! This would make it one of, if not the, largest known dinosaurs - and certainly the longest! Though it does have a long tail, it differs in appearance from its cousin Diplodocus primarily by having a proportionally longer neck and shorter tail. It was also more slender than Apatosaurus, though it was longer than that contemporary. How did Barosaurus get such a long neck? It literally converted one of the back vertebrae into a neck vertebra! This is so fascinating that I can’t get over it - its close relatives, like Diplodocus, did not employ this to get a longer neck, indicating Barosaurus was using its long neck for things that its cousins were not. Barosaurus was also weird in not having as high of spines on its vertebrae as its cousin Diplodocus and other members of the group. In addition to all of that - it had shorter vertebrae in the tail, which made it shorter than in other members of this group! Interestingly, the bones on the underside of the tail were forked and had forward spikes, which would have given it similar strength to that of Diplodocus; it was probably still a whiptail like other members of this group, though not as much of one as its relatives.

By Slate Weasel, in the Public Domain
Of course, the distinctiveness of Barosaurus is primarily limited to the length of the animal and its spine. In terms of limbs, it had fairly identical limbs to its cousin Diplodocus, though it did have fairly long forelimbs compared to its cousin (by… an almost imperceptible amount, however). Though the feet of Barosaurus aren’t known, it is reasonable to suppose that it would have had feet similar to Diplodocus - with only one claw on the front feet and three small claws on the hind feet. The skull of Barosaurus is not known, but it probably would have been long and low, with peg-like teeth in the front of the jaws for grazing on plants. Its neck was not very flexible in the vertical sense, but it was much more flexible in sweeping from side to side. It is possible that there were spikes of some sort at the end of the tail, which would have packed quite a punch when the tail was used to whip other animals. And, finally, it would have been entirely - if not almost entirely - scaly all over its body. It is also possible that Barosaurus may have featured some brilliant colors, especially in the tail, for communication with other members of the species.
Diet: Barosaurus would have primarily fed on high-level vegetation, able to reach much of it at its natural neck height and then - on top of that - being able to rear up to 50 meters high via going on its hind legs. However, a lack of vertical reach in terms of neck flexibility means that it probably would have swept over a wide area for food, rather than going up and down in the tree level like other Diplodocids. This would have allowed Barosaurus to move very little - if at all - while eating, instead of moving over large distances in search of vegetation.

By Scott Reid
Behavior: Barosaurus was not especially common in its environment, so the question of its social nature is actually somewhat important. Fossil evidence indicates at least some sociality in other Diplodocids - herding, or at least small herds, of other sauropods on the Morrison are clear from fossil evidence and trackways. The question remains - did Barosaurus do what its cousins did? The question is, of course, up in the air without more fossil evidence. It is possible that, in an environment with hundreds and hundreds of large sauropods to feed, Barosaurus may have been more solitary to aid in getting enough food without competing too much with one another. Alternatively, it may have also lived in social groups, allowing for the safety of weaker members of the herd and more cohesiveness in finding food.
Barosaurus, like other Diplodocids, would have been able to rear up on its hind legs to get food. This action would have also made Barosaurus even taller than usual, which would have been fairly imposing to predators nearby. It had a whip-tail, which would have allowed Barosaurus to make very loud sonic cracks in the air; if that tail was covered with spikes, as in other members of the group, it would have also lacerated the skin of other dinosaurs. Still, even without spikes, it would have packed quite a punch for any predators that might have tried to attack it. The sounds of the tail would have been a warning; it is possible that such sounds would have been used in communication with one another, and potentially even display in competition for mates and food and similar things. The impossibly long neck probably was also a sort of sexual display structure, since the longer neck indicated being able to reach more food without walking around. It is uncertain whether or not it would have taken care of its young; while there is no evidence either way - which usually would lead to concluding it did, given the fact most living archosaurs do and there’s extensive evidence of such in extinct dinosaurs - other sauropods (aka the titanosaurs) probably didn’t. So, for now, the jury on that is out.

By Fred Wierum, CC BY-SA 4.0
Ecosystem: Barosaurus lived in the Morrison Formation - an extensive, expansive semi-arid, seasonal floodplain that covered most of Western North America during the Jurassic and was filled with iconic dinosaurs and other animals that we usually think of when we think of the “Jurassic.” Though the Morrison was as arid and open as a modern savanna, the lack of extensive flowering plants at this time rendered the habitat more like a ridiculously huge scrubland. There were a variety of trees - conifers, ginkgos, cycads, and tree ferns - dispersed among the bushes and horsetails and other plants. They congregated around rivers, which were havens of life amongst the arid territory. At the time of the Brushy Basin Environment - the last part of the formation, where Barosaurus could be found - this environment was much muddier and wetter, potentially indicating a change in ecology that would lead to the end of the Morrison Formation, and an extinction of the animals there. There were also expansive volcanic explosions that lead to much of the preservation we see there. A large salt lake present would have been a major feature of the environment, and it was connected to extensive wetlands that formed a break in the wider scrubland around the habitat.

By Danny Cicchetti, CC BY-SA 3.0
Barosaurus may be known from the entire Brushy Basin Environment of the Morrison; however, confirmed fossils of this dinosaur are only known from a few sites. So, in my map above, I give two colors - the wider green color to show the whole ecosystem, aka the wider area that Barosaurus may have ventured in to; and the smaller blue color to show the confirmed range of this dinosaur. In that confirmed range, Barosaurus lived alongside a lot of other animals - in fact, there’s a reason the Morrison is so iconic - its characteristic and distinctive fossils, both of dinosaurs and not of dinosaurs. Barosaurus has been found in, literally, the same sites as other animals - it is known to have lived alongside the predator Allosaurus; in another site, turtles and Pseudosuchians and the Choristodere Cteniogenys, as well as Allosaurus and the more bulky sauropod Camarasaurus; in yet another, Barosaurus lived alongside many turtles, the Pseudosuchians Hoplosuchus and Goniopholis, the tuatara-like Opisthias, and a wide variety of dinosaurs - other sauropods like Diplodocus Apatosaurus and Camarasaurus, predators like Allosaurus Torvosaurus and Ceratosaurus, and Ornithiscians like Stegosaurus Dryosaurus and Uteodon. So, Barosaurus was a part of a very wide and diverse community, with a great diversity in terms of herbivores and predators that would have attacked Barosaurus. That being said, there were many other animals that may have lived alongside Barosaurus, based on just… probability, even though they weren’t found directly with it. There were other stegosaurs like Alcovasaurus and Hesperosaurus; more small running herbivores like Nanosaurus; larger bulky bipedal herbivores like Camptosaurus; more sauropods, including Apatosaurus and Supersaurus; and smaller predators that would have probably been more of a threat to Barosaurus young than adults - Marshosaurus, Coelurus, Ornitholestes, and Stokesosaurus. Sadly, the organization of the Morrison is something of a mess - so, while many other dinosaurs and animals lived alongside Barosaurus, we can’t exactly be sure which ones. There were probably a variety of Multituberculate, Tinodontid, Eutriconodont, and Dryolestoid mammals, as well as others; some pterosaurs were probably there like Harpactognathus, and, of course, there were amphibians as well. This makes the Morrison one of the better examples of an environment to highlight as a representative of a particular time in Earth’s history - since it showcases so many different living things!

By José Carlos Cortés
Other: Barosaurus was a close relative of Diplodocus, though it is difficult to determine how close, as the evolutionary relationships between the Diplodocids are still being worked out via phylogenetic studies. It is possible that an offshoot of Diplodocus (which were around before Barosaurus evolved) split to take advantage of not moving much to eat, and instead sweeping its neck around to gather food. For a while, another sauropod in Africa was considered to be a species of Barosaurus; today, however, it seems to be very clearly in its own genus, Tornieria, and actually far removed from both Diplodocus and Barosaurus (while still being in this closely related family group). So, for now, Barosaurus is only known from North America. These dinosaurs were distinctive long and slender sauropods, as opposed to their long and bulky cousins, the Apatosaurines, that they lived alongside.
~ By Meig Dickson
Sources Under the Cut
Allen, Eric Randall (Summer 2012). "Analysis of North American goniopholidid crocodyliforms in a phylogenetic context.” University of Iowa Research Online.
Apesteguía, S. 2005. Evolution in the hyposphene-hypantrum complex within Sauropoda. In K. Carpenter and V. Tidwell (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press, Bloomington 248-267.
Arldt, T. 1909. Die Dinosaurier [The dinosaurs]. Naturwissenshaftliche Rundschau 24(21):261-263.
Averianov, A. O., and T. Martin (2015). "Ontogeny and taxonomy of Paurodon valens (Mammalia, Cladotheria) from the Upper Jurassic Morrison Formation of USA" (PDF). Proceedings of the Zoological Institute of the Russian Academy of Sciences. 319 (3): 326–340.
Berman, D. S., and J. S. McIntosh. 1978. Skull and relationships of the Upper Jurassic sauropod Apatosaurus (Reptilia, Saurischia). Bulletin of Carnegie Museum of Natural History 8:1-35.
Bilbey, S.A. (1998). "Cleveland-Lloyd Dinosaur Quarry - age, stratigraphy and depositional environments". In Carpenter, K.; Chure, D.; and Kirkland, J.I. (eds.) (eds.). The Morrison Formation: An Interdisciplinary Study. Modern Geology 22. Taylor and Francis Group. pp. 87–120.
Bonaparte, J. F. 1986. The early radiation and phylogenetic relationships of the Jurassic sauropod dinosaurs, based on vertebral anatomy. In K. Padian (ed.), The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic–Jurassic Boundary. Cambridge University Press, Cambridge 247-258
Bonaparte, J. F. 1986. Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétosauridés) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentina) [The dinosaurs (carnosaurs, allosaurids, sauropods, cetiosaurids) from the Middle Jurassic of Cerro Cóndor (Chubut, Argentina)]. Annales de Paléontologie (Vert.-Invert.) 72(3):325-386.
Britt, B. B., and B. G. Naylor. 1994. An embryonic Camarasaurus (Dinosauria, Sauropoda) from the Upper Jurassic Morrison Formation (Dry Mesa Quarry, Colorado). In K. Carpenter, K. F. Hirsch, and J. R. Horner (eds.), Dinosaur Eggs and Babies 256-264.
Butler, R.J., P.M. Galton, L.B. Porro, L.M. Chiappe, D.M. Henderson, and G.M. Erickson. 2009. Lower limits of ornithischian dinosaur body size inferred from a new Upper Jurassic heterodontosaurid from North America. Proceedings of the Royal Society B 10.1098/rspb.2009.1494.
Caldwell, M. W.; Nydam, R. L.; Palci, A.; Apesteguía, S. N. (2015). "The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution". Nature Communications. 6: 5996.
Calvo, J. O., and L. Salgado. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. GAIA 11:13-33.
Carpenter K & Galton PM (2001). "Othniel Charles Marsh and the Eight-Spiked Stegosaurus". In Carpenter, Kenneth (ed.). The Armored Dinosaurs. Indiana University Press. pp. 76–102.
Carpenter, Kenneth (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus". In Foster, John R.; Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131–138.
Carpenter, K. and Wilson, Y. 2008. A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb. Annals of the Carnegie Museum 76:227-263.
Carpenter, Kenneth (2018). Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado. Geology of the Intermountain West. 5: 227–244.
Carrano and Sampson, 2008. The phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 6, 183-236.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution 1-698
Charig, A. J. 1980. A diplodocid sauropod from the Lower Cretaceous of England. In L. L. Jacobs (ed.), Aspects of Vertebrate History: Essays in Honor of Edwin Harris Colbert. Museum of Northern Arizona Press, Flagstaff 231-244
Chure, 2001. The second record of the African theropod Elaphrosaurus (Dinosauria, Ceratosauria) from the Western Hemisphere. Neues Jahrbuch für Geologie und Paläontologie Monatshefte. 2001(9).
Chure, Daniel J. (2001). "On the type and referred material of Laelaps trihedrodon Cope 1877 (Dinosauria: Theropoda)". In Tanke, Darren; and Carpenter, Kenneth (eds.) (eds.). Mesozoic Vertebrate Life. Bloomington and Indianapolis: Indiana University Press. pp. 10–18.
Chure, D. J., R. Litwin, S. T. Hasiotis, E. Evanoff, and K. Carpenter. 2006. The fauna and flora of the Morrison Formation: 2006. In J. R. Foster, S. G. Lucas (eds.), Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36:233-249
Coombs, W. P., and R. E. Molnar. 1981. Sauropoda (Reptilia, Saurischia) from the Cretaceous of Queensland. Memoirs of the Queensland Museum 20(2):351-373
Curtice, B. D. 1995. A description of the anterior caudal vertebrae of Supersaurus vivianae. Journal of Vertebrate Paleontology 15(3, suppl.):25A
Dalman, S.G. (2014). "New data on small theropod dinosaurs from the Upper Jurassic Morrison Formation of Como Bluff, Wyoming, USA". Volumina Jurassica. 12 (2): 181–196.
Demko, Timothy M.; Parrish, Judith T. (1998). "Paleoclimatic setting of the Upper Jurassic Morrison Formation". In Carpenter, Ken; Chure, Daniel J.; Kirkland, James I. (eds.). The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology 22 (1-4): 283-296.
Dodson, P. 1997. American dinosaurs. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 10-13
Ekart, Douglas D.; Cerling, Thure E.; Montanez, Isabel P.; Tabor, Neil J. (1999). "A 400 million year carbon isotope record of pedogenic carbonate; implications for paleoatmospheric carbon dioxide" (PDF). American Journal of Science. 299 (10): 805–827.
Engelmann, George F.; Chure, Daniel J.; Fiorillo, Anthony R. (2004). "The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation". In Turner, Christine E.; Peterson, Fred; Dunagan, Stan P. (eds.). Reconstruction of the Extinct Ecosystem of the Upper Jurassic Morrison Formation. Sedimentary Geology. Sedimentary Geology 167 (3-4): 297-308. 167. pp. 297–308.
Foster, John R. (1996). "Sauropod dinosaurs of the Morrison Formation (Upper Jurassic), Black Hills, South Dakota and Wyoming". Contributions to Geology, University of Wyoming. 31 (1): 1–25.
Foster, J.R. 2003. Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. Bulletin 23.
Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327-329.
Foster, J. 2007. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 389pp.
Foster, J.R. 2009. Preliminary body mass estimates for mammalian genera of the Morrison Formation (Upper Jurassic, North America). PaleoBios 28(3):114-122.
Foster, J. (2018). "A new atoposaurid crocodylomorph from the Morrison Formation (Upper Jurassic) of Wyoming, USA". Geology of the Intermountain West. 5: 287–295.
Fraas, Eberhard (1908). "Ostafrikanische Dinosaurier". Palaeontographica. 55: 105–144.
Galianom, H., and R. Albersdörfer. 2010. A New Basal Diplodocoid Species, Amphicoelias brontodiplodocus from the Morrison Formation, Big Horn Basin, Wyoming, with Taxonomic Reevaluation of Diplodocus, Apatosaurus, Barosaurus and Other Genera. Dinosauria International (Ten Sleep, WY) Report for September 2010 1-41
Gallina, P. A., and S. Apesteguía. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista del Museo Argentino de Ciencias Naturales, nuevo serie 7(2):153-166
Galton, P. M. 1977. The Upper Jurassic dinosaur Dryosaurus and a Laurasia-Gondwana connection in the Upper Jurassic. Nature 268(5617):230-232
Galton, P.M. & Powell, H.P. (1980). "The ornithischian dinosaur Camptosaurus prestwichii from the Upper Jurassic of England". Palaeontology. 23: 411–443.
Galton, P.M. (1981). Dryosaurus, a hypsilophodontid dinosaur from the Upper Jurassic of North America and Africa. Postcranial skeleton. Palaeontol. Z. 55(3/4), 271-312
Galton, 1982. Elaphrosaurus, an ornithomimid dinosaur from the Upper Jurassic of North America and Africa. Paläontologische Zeitschrift. 56, 265-275.
Galton PM, Upchurch P (2004). "Stegosauria". In Weishampel DB, Dodson P, Osmólska H. The Dinosauria (2nd Edition). University of California Press. p. 361.
Galton, P.M. (2010). "Species of plated dinosaur Stegosaurus (Morrison Formation, Late Jurassic) of western USA: new type species designation needed". Swiss Journal of Geosciences 103 (2): 187–198.
Galton, Peter M. & Carpenter, Kenneth, 2016, "The plated dinosaur Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; Upper Jurassic, western USA), type species of Alcovasaurus n. gen.", Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 279(2): 185-208.
Gillette, D.D. (1991). "Seismosaurus halli, gen. et sp. nov., a new sauropod dinosaur from the Morrison Formation (Upper Jurassic/Lower Cretaceous) of New Mexico, USA". Journal of Vertebrate Paleontology. 11 (4): 417–433.
Gilmore, C.W. (1909). "Osteology of the Jurassic reptile Camptosaurus, with a revision of the species of the genus, and descriptions of two new species". Proceedings of the United States National Museum. 36 (1666): 197–332.
Harris, J. D., and P. Dodson. 2004. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontologica Polonica 49(2):197-210
Harris, J. D. 2006. The axial skeleton of the dinosaur Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Palaeontology 49(5):1091-1121.
Hartman, S.; Mickey Mortimer; William R. Wahl; Dean R. Lomax; Jessica Lippincott; David M. Lovelace (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247.
Haughton, S. H. 1928. On some reptilian remains from the Dinosaur Beds of Nyasaland. Transactions of the Royal Society of South Africa 16:67-75
Hay, O. P. 1902. Bibliography and Catalogue of the Fossil Vertebrata of North America. Bulletin of the United States Geological Survey 179:1-868
Hendrickx, C, Mateus O. 2014. Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods, 03. PLoS ONE. 9:e88905., Number 3.
Holtz, Thomas R., Jr.; Molnar, Ralph E.; Currie, Philip J. (2004). Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 71–110.
Huene, F. v. 1908. Die Dinosaurier der Europäischen Triasformation mit berücksichtigung der Ausseuropäischen vorkommnisse [The dinosaurs of the European Triassic formations with consideration of occurrences outside Europe]. Geologische und Palaeontologische Abhandlungen Suppl. 1(1):1-419
Huene, F. v. 1909. Skizze zu einer Systematik und Stammesgeschichte der Dinosaurier [Sketch of the systematics and origins of the dinosaurs]. Centralblatt für Mineralogie, Geologie und Paläontologie 1909:12-22
Huene, F. v. 1927. Short review of the present knowledge of the Sauropoda. Memoirs of the Queensland Museum 9(1):121-126
Huene, F. v. 1927. Sichtung der Grundlagen der jetzigen Kenntnis der Sauropoden [Sorting through the basis of the current knowledge of sauropods]. Eclogae Geologica Helveticae 20:444-470
Huene, F. v. 1929. Los sauriquios y ornitisquios del Cretáceo argentino. Anales del Museo de La Plata, serie 2 3:1-196
Ibiricu, L. M., G. A. Casal, M. C. Lamanna, R. D. Martínez, J. D. Harris and K. J. Lacovara. 2012. The southernmost records of Rebbachisauridae (Sauropoda: Diplodocoidea), from early Late Cretaceous deposits in central Patagonia. Cretaceous Research 34:220-232
Janensch, W. 1914. Übersicht über die Wirbeltierfauna der Tendaguru-Schichten [Overview of the vertebrate fauna of the Tendaguru beds]. Archiv für Biontologie 3:81-110
Janensch, Werner (1922). "Das Handskelett von Gigantosaurus robustus und Brachiosaurus brancai aus den Tendaguru-Schichten Deutsch-Ostafrikas". Centralblatt für Mineralogie, Geologie und Paläontologie. 1922: 464–480.
Jenkins, J.T. and J.L. Jenkins. 1993. Colorado's Dinosaurs. Denver, Colorado: Colorado Geologic Survey. Special Publication 35.
Joleaud, L. 1922. Les reptiles fossiles [Fossil reptiles]. Association Française pour l'Avancement des Sciences. Conférences. Compte Rendu de la 45e Session 49-66
Kenneth Carpenter; Peter M. Galton (2018). "A photo documentation of bipedal ornithischian dinosaurs from the Upper Jurassic Morrison Formation, USA". Geology of the Intermountain West. 5: 167–207.
Kirkland, J. I. 1997. Cedar Mountain Formation. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 98-99.
Kowallis, Bart J.; Christiansen, Eric H.; Deino, Alan L.; Peterson, Fred; Turner, Christine E.; Kunk, Michael J.; Obradovich, John D. (1998). "The age of the Morrison Formation" (PDF). In Carpenter, Ken; Chure, Daniel J.; Kirkland, James I. (eds.). The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology 22 (1-4): 235-260.
Ksepka, D. T., and M. A. Norell. 2010. The Illusory Evidence for Asian Brachiosauridae: New Material of Erketu ellisoni and a Phylogenetic Reappraisal of Basal Titanosauriformes. American Museum Novitates 3700:1-27.
Lockley, M.; Harris, J.D.; and Mitchell, L. 2008. "A global overview of pterosaur ichnology: tracksite distribution in space and time." Zitteliana. B28. p. 187-198.
Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional, Rio de Janeiro. 65 (4): 527–544.
Lucas S, Herne M, Heckert A, Hunt A, and Sullivan R. Reappraisal of Seismosaurus, A Late Jurassic Sauropod Dinosaur from New Mexico. The Geological Society of America, 2004 Denver Annual Meeting (7–10 November 2004).
Lucas, Spencer G.; Spielmann, Justin A.; Rinehart, Larry A.; Heckert, Andrew B; Herne, Matthew C.; Hunt, Adrian P.; Foster, John R.; Sullivan, Robert M. (2006). "Taxonomic status of Seismosaurus hallorum, a Late Jurassic sauropod dinosaur from New Mexico". In Foster, John R.; Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 149-161.
Lull, R. S. 1919. The sauropod dinosaur Barosaurus Marsh: redescription of the type specimens in the Peabody Museum, Yale University. Memoirs of the Connecticut Academy of Arts and Sciences 6:1-42.
Lull, R. S. 1924. Dinosaurian climatic response. In M. R. Thorpe (ed.), Organic Adaptation to Environment 225-279.
Lydam, R. L., Daniel J. Chure and Susan E. Evans (2013). "Schillerosaurus gen. nov., a replacement name for the lizard genus Schilleria Evans and Chure, 1999 a junior homonym of Schilleria Dahl, 1907" (PDF). Zootaxa. 3734 (1): 99–100.
Madsen, J. H., and W. E. Miller. 1979. the fossil vertebrates of Utah, an annotated bibliography. Brigham Young University Geology Studies 26(4):iii-147
Maidment, Susannah C.R.; Norman, David B.; Barrett, Paul M.; Upchurch, Paul (2008). "Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia)". Journal of Systematic Palaeontology. 6 (4): 367–407.
Mannion, P. D., P. Upchurch, O. Mateus, R. N. Barnes, and M. E. H. Jones. 2012. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology 10(3):521-551
Marsh, O. C. 1890. Description of new dinosaurian reptiles. The American Journal of Science, series 3 39:81-86.
Marsh, O. C. 1895. On the affinities and classification of the dinosaurian reptiles. American Journal of Science 50(300):483-498.
Marsh, O. C. 1896. The dinosaurs of North America. United States Geological Survey, 16th Annual Report, 1894-95 55:133-244.
Marsh, O. C. 1898. On the families of sauropodous Dinosauria. Geological Magazine, decade 4 3:157-158.
Marsh, Othniel C. (1899). "Footprints of Jurassic dinosaurs". American Journal of Science. 4 (7): 227–232.
Martin, John. 1987. Mobility and feeding of Cetiosaurus (Saurischia, Sauropoda) – why the long neck? pp. 154–159 in P. J. Currie and E. H. Koster (eds), Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Boxtree Books, Drumheller (Alberta). 239 pages.
Mateus, O., & Antunes M. T. 2000. Ceratosaurus sp. (Dinosauria: Theropoda) in the Late Jurassic of Portugal. Abstract volume of the 31st International Geological Congress, Rio de Janeiro, Brazil.
Mateus, O. 2006. Late Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): a comparison. In J. R. Foster, S. G. Lucas (eds.), Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36:223-231.
Mateus, O. (2007). Notes and review of the ornithischian dinosaurs of Portugal. Journal of Vertebrate Paleontology. 27, 114A-114A., Jan: Society of Vertebrate Paleontology.
Mateus, O., & Tschopp E. (2013). Cathetosaurus as a valid sauropod genus and comparisons with Camarasaurus. Journal of Vertebrate Paleontology, Program and Abstracts, 2013. 173.
McIntosh, J. S. 1981. Annotated catalogue of the dinosaurs (Reptilia, Archosauria) in the collections of the Carnegie Museum of Natural History. Bulletin of Carnegie Museum of Natural History 18:1-67.
McIntosh, J. S. 1990. Sauropoda. In D. B. Weishampel, H. Osmólska, and P. Dodson (eds.), The Dinosauria. University of California Press, Berkeley 345-401.
McIntosh, J. S. 1997. Sauropoda. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 654-658.
McIntosh, J. S. 2005. The genus Barosaurus Marsh (Sauropoda, Diplodocidae). In K. Carpenter and V. Tidwell (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press, Bloomington 38-77.
Melstrom, K. M., M. D. D'Emic, D. J. Chure and J. A. Wilson. 2016. A juvenile sauropod dinosaur from the Late Jurassic of Utah, U.S.A., presents further evidence of an avian style air-sac system. Journal of Vertebrate Paleontology 36(4):e1111898:1-23.
Mezga, A., B. C. Tesovic, and Z. Bajraktarevic. 2007. First record of dinosaurs in the Late Jurassic of the Adriatic-Dinaridic carbonate platform (Croatia). Palaios 22(2):188-199.
Miller, W. E., J. L. Baer, K. L. Stadtman and B. B. Britt. 1991. The Dry Mesa Dinosaur Quarry, Mesa County, Colorado. In W. R. Averett (ed.), Guidebook for Dinosaur Quarries and Tracksites Tour, Western Colorado and Eastern Utah 31-46.
Mocho, P., R. Royo-Torres, and F. Ortega. 2014. Phylogenetic reassessment of Lourinhasaurus alenquerensis, a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal. Zoological Journal of the Linnean Society 170:875-916.
Moore, George T.; Hayashida, Darryl N.; Ross, Charles A.; Jacobson, Stephen R. (1992). "Paleoclimate of the Kimmeridgian/Tithonian (Late Jurassic) world: I. Results using a general circulation model". Palaeogeography, Palaeoclimatology, Palaeoecology. 93 (3–4): 113–150.
Nopcsa, B. F. 1928. The genera of reptiles. Palaeobiologica 1:163-188.
Olshevsky, G. 1992. A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia. Mesozoic Meanderings 2:1-268.
Parrish, J.T.; Peterson, F.; Turner, C.E. (2004). "Jurassic "savannah"-plant taphonomy and climate of the Morrison Formation (Upper Jurassic, Western USA)". Sedimentary Geology. 167 (3–4): 137–162.
Pickering, 1995a. Jurassic Park: Unauthorized Jewish Fractals in Philopatry. A Fractal Scaling in Dinosaurology Project, 2nd revised printing. Capitola, California. 478 pp.
Pritchard, A. C.; Turner, A. H.; Allen, E. R.; Norell, M. A. (2013). "Osteology of a North American Goniopholidid (Eutretauranosuchus delfsi) and Palate Evolution in Neosuchia". American Museum Novitates 3783 (3783).
Rauhut, O. W. M., K. Remes, R. Fechner, G. Cladera, and P. Puerta. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435:670-672.
Remes, K. 2006. A revision of the Tendaguru sauropod dinosaur Tornieria africana (Fraas) and its relevance for sauropod paleobiogeography. Journal of Vertebrate Paleontology 26(3):651-669.
Remes, K. 2007. A second Gondwanan diplodocid dinosaur from the Upper Jurassic Tendaguru Beds of Tanzania, East Africa. Palaeontology 50(3):653-667.
Romer, A. S. 1956. Osteology of the Reptiles, University of Chicago Press 1-77.
Romer, A. S. 1966. Vertebrate Paleontology, 3rd edition 1-468.
Royo-Torres, R., A. Cobos, A. Aberasturi, E. Espílez, I. Fierro, A. González, L. Luque, L. Mampel, and L. Alcalá. 2007. Riodeva sites (Teruel, Spain) shedding light to European sauropod phylogeny. Geogaceta 41:183-186.
Ruiz-Omeñaca, J. I., L. Piñuela, and J. C. García-Ramos. 2008. Primera evidencia de dinosaurios diplodocinos (Sauropoda: Diplodocidae) en el Jurásico Superior de Asturias (Noreña) [First evidence of diplodocine dinosaurs (Sauropoda: Diplodocidae) in the Upper Jurassic of Asturias (Noreña)]. In J I Ruiz-Omeñaca, L Piñuela and J C García-Ramos (eds), XXIV Jornadas de la Sociedad Española de Paleontología, 15-18 October 2008, Museo del Jurásico de Asturias (MUJA), Colunga, Spain, Libro de Resúmenes 191-192.
Russell, D., P. Beland, and J. McIntosh. 1980. Paleoecology of the dinosaurs of Tendaguru (Tanzania). Mem. Society Geol. France, N.S. 139:169-175.
Russell, D. A. 1984. A check list of the families and genera of North American dinosaurs. Syllogeus 53:1-35.
Saleiro, A., & Mateus O. (2017). Upper Jurassic bonebeds around Ten Sleep, Wyoming, USA: overview and stratigraphy. Abstract book of the XV Encuentro de Jóvenes Investigadores en Paleontología/XV Encontro de Jovens Investigadores em Paleontologia, Lisboa, 428 pp.. 357-361.
Salgado, L. 1993. Comments on Chubutisaurus insignis del Corro (Saurischia, Sauropoda). Ameghiniana 30(3):265-270.
Salgado, L. 1999. The macroevolution of the Diplodocimorpha (Dinosauria; Sauropoda): a developmental model. Ameghiniana 36(2):203-216.
Salgado, L., I. d. S. Carvalho, and A. C. Garrido. 2006. Zapalasaurus bonapartei, un nuevo saurópodo de La Formación La Amarga (Cretacico Inferior), noroeste de Patagonia, Provincia de Neuquén, Argentina [Zapalasaurus bonapartei, a new sauropod from the La Amarga Formation (Lower Cretaceous), northwestern Patagonia, Neuquén province, Argentina]. Géobios 39:695-707.
Schultz, J. A; Bhart-Anjan S. Bhullar; Zhe-Xi Luo (2018). "Re-examination of the Jurassic mammaliaform Docodon victor by computed tomography and occlusal functional analysis". Journal of Mammalian Evolution. in press. doi:10.1007/s10914-017-9418-5.
Seebacher, Frank. (2001). "A new method to calculate allometric length-mass relationships of dinosaurs". Journal of Vertebrate Paleontology. 21 (1): 51–60.
Seeley, Harry G. (1869). Index to the fossil remains of Aves, Ornithosauria and Reptilia, from the Secondary system of strata arranged in the Woodwardian Museum of the University of Cambridge. Cambridge: Deighton, Bell and Co. pp. 143pp.
Sereno, P. C. 1997. The origin and evolution of dinosaurs. Annual Review of Earth and Planetary Sciences 25:435-489.
Smith, David K. (1998). "A morphometric analysis of Allosaurus". Journal of Vertebrate Paleontology. 18 (1): 126–142.
Smith, D. M., M. A. Gorman, J. D. Pardo and B. J. Small. 2011. First fossil Orthoptera from the Jurassic of North America. Journal of Paleontology 85(1):102-105.
Steel, R. 1970. Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology. Gustav Fischer Verlag, Stuttgart 1-87.
Sternfeld, R. 1911. Zur Nomenklatur der Gattung Gigantosaurus Fraas [On the nomenclature of the genus Gigantosaurus Fraas]. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin 8:398.
Suteethorn, S., J. Le Loeuff, E. Buffetaut and V. Suteethorn. 2010. Description of topotypes of Phuwiangosaurus sirindhornae, a sauropod from the Sao Khua Formation (Early Cretaceous) of Thailand, and their phylogenetic implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 256(1):109-121.
Swinton, W. E. 1970. The Dinosaurs, Wiley-Interscience, New York 1-331.
Tatarinov, L. P. 1964. Nadotryad Dinosauria. Dinozavry [Superorder Dinosauria. Dinosaurs]. In Y. A. Orlov (ed.), Osnovy Paleontologii [Fundamentals of Paleontology] 12:523-589.
Taylor, Michael P; Wedel, Mathew J (2013). "The neck of Barosaurus was not only longer but also wider than those of Diplodocus and other diplodocines". PeerJ. 1: e67v1.
Tidwell, V., Carpenter, K., and Miles, C., 2005, A reexamination of Morosaurus agilis (Sauropoda) from Garden Park, Colorado: Journal of Vertebrate Paleontology 25 (Supplement to 3):122A.
Tornier, G. 1913. Reptilia. Paläontologie [Reptilia. Paleontology]. Handwörterbuch der Naturwissenschaften 8:337-376.
Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. (2006). "Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations". Geological Society of America Abstracts with Programs. 38 (6): 7.
Tschopp, E., and O. Mateus. 2013. The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs. Journal of Systematic Palaeontology 11(7):853-88.
Tschopp, E., O. Mateus, and R. B. J. Benson. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 3:e857.
Turner, Christine E.; Peterson, Fred (2004). "Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis". In Turner, Christine E.; Peterson, Fred; Dunagan, Stan P. (eds.). Reconstruction of the Extinct Ecosystem of the Upper Jurassic Morrison Formation. Sedimentary Geology. Sedimentary Geology 167 (3-4): 309-355.
Upchurch, P. 1995. The evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London B 349:365-390.
Upchurch, P. 1995. Sauropod phylogeny and palaeoecology. In M. G. Lockley, V. F. dos Santos, C. A. Meyer, & A. P. Hunt (eds.), Aspects of Sauropod Paleobiology. GAIA 10:249-260.
Upchurch, P., P. M. Barrett, and P. Dodson. 2004. Sauropoda. In D. B. Weishampel, H. Osmolska, and P. Dodson (eds.), The Dinosauria (2nd edition). University of California Press, Berkeley 259-322.
von Zittel, K. A. v. 1890. Handbuch der Palaeontologie. I. Abteilung Paleozoologie. III. Band. Vertebrata (Pisces, Amphibia, Reptilia, Aves) [Handbook of Paleontology. Division I. Paleozoology. Volume III. Vertebrata (Pisces, Amphibia, Reptilia, Aves)] xii-900.
von Zittel, K. A. 1911. Grundzüge der Paläontologie (Paläozoologie). II. Abteilung. Vertebrata [Fundamentals of Paleontology (Paleozoology). Section II. Vertebrata]. Druck und Verlag von R. Oldenbourg, München 1-598.
Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. 861 pp.
Whitlock, J. A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161:872-915.
Wieland, G. R. 1920. The longneck sauropod Barosaurus. Science, New Series 51(1326):528-530.
Wild, Rupert (1991). "Janenschia n. g. robusta (E. Fraas 1908) pro Tornieria robusta (E. Fraas 1908) (Reptilia, Saurischia, Sauropodomorpha)". Stuttgarter Beiträge zur Naturkunde, Serie B: Geologie und Paläontologie. 173: 1–4.
Wilson, J. A., and M. B. Smith. 1996. New remains of Amphicoelias Cope (Dinosauria: Sauropoda) from the Upper Jurassic of Montana and diplodocoid phylogeny. Journal of Vertebrate Paleontology 16(3, suppl.):73A.
Wilson, J. A., and P. C. Sereno. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5. Journal of Vertebrate Paleontology 18(2 (suppl.)):1-68.
Wilson, J. A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136:217-276.
Xing, L., T. Miyashita, J. Zhang, D. Li, Y. Te, T. Sekiya, F. Wang and P. J. Currie. 2015. A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids. Journal of Vertebrate Paleontology 35(1):e889701:1-17.
#Barosaurus lentus#Barosaurus#Dinosaur#Diplodocid#Diplodocoid#Factfile#Palaeoblr#Jurassic#Herbivore#North America#Mesozoic Monday#paleontology#prehistory#prehistoric life#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#Sauropod
332 notes
·
View notes
Text
Xingxiulong chengi
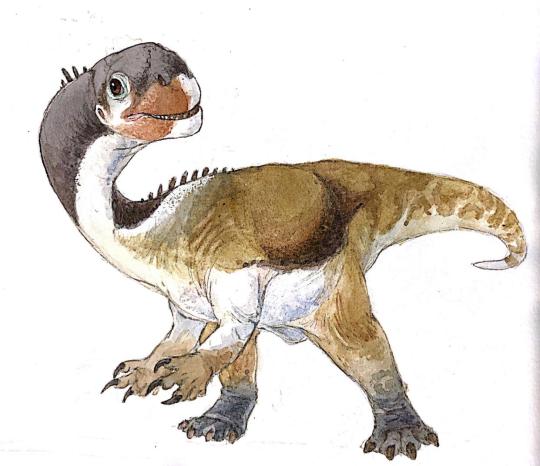
By Ripley Cook
Etymology: Dragon from the Xingxiu Bridge
First Described By: Wang et al., 2017
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes
Status: Extinct
Time and Place: Between 201.3 and 199.3 million years ago, in the Hettangian of the Early Jurassic


Xingxiulong is known from the Shawan Member of the Lufeng Formation

Physical Description: Xingxiulong was a medium-sized prosauropod, about four to five meters long with a height of one to one and a half meters at the hip. It had a smooth jaw, unlike other prosauropods of the time and place, and it had a long jaw like Lufengosaurus. It’s skull was somewhat short, much like that of the later Camarasaurus, and it would have relied more on its neck and hands to grab food rather than a longer snout. This broad snout would have also made it a generalist, rather than specialist, browser. It’s neck was fairly long and muscular, like in other Sauropodomorphs; it also had a long tail.
Xingxiulong was a fairly bulky and heavyset prosauropod, indicating the journey of this line of dinosaurs as it evolved into Sauropods (though of course, Sauropods were already around at this point in dinosaurian evolution). It even had more vertebrae at the hip, like sauropods, and unlike other prosauropods of its time and place. It also had a longer hip bone than prosauropods - once again, one more like the later sauropods. It’s legs were also very robust, like those of the sauropods. Xingxiulong had a very large gut, allowing it to store more food while digesting. This probably is correlated with its increased ability to hold up a large body weight, compared to its contemporaries. In short, Xingxiulong is a prime example of the slow, step-wise evolution of the prosauropods towards the large nature of the Sauropods - though,of course, Xingxiulong is just an offshoot of that line.
Being somewhat larger, Xingxiulong probably was more scaly than not, though it still may have possessed feathers. It probably would have stille mostly walked on just its hindlimbs, though it may have been able to walk on its forelimbs when need be. It still would have mainly used its forelimbs in feeding, more than in locomotion.

By José Carlos Cortés
Diet: Xingxiulong would have been a mid-level browser, feeding on a variety of fairly dry vegetation by reaching up with its arms and long neck.
Behavior: Xingxiulong, unfortunately, doesn’t have a very well-known behavior, though it probably would have spent most of its time browsing in its environment. It would have had to walk around its environment carefully, given how bulky it was and its large mass; it probably wouldn’t have been the fastest of dinosaurs. As a prosauropod, it most likely was active in its metabolism, and it probably took care of its young. It’s also entirely likely that Xingxiulong would forage in its environment in mixed-species herds, with all the other prosauropods of the time and place in which it lived.
Ecosystem: Xingxiulong lived in the Shawan environment of the Lufeng Formation, one of a few classic environments showing off the initial explosion of dinosaur evolution after the end-Triassic extinction. This was a lush environment, with many lakes, rivers, and overbanks. As such, it would have been a very lush and green environment, though no plant fossils have been reported. Here there were many other kinds of animals, especially transitional dinosaurs. In many ways, it was a counterpart to the South African Elliot Formation, in what it shows about the early evolution of large dinosaurs - though the Elliot was much more arid than the Lufeng.

By Scott Reid
Contemporary with Xingxiulong, there were many other Prosauropods that probably would have all grazed together in the plant line in mixed-species herds. Here, there was Jingshanosaurus, Yunnanosaurus, Lufengosaurus, and Gyposaurus, all in the specific environment from which Xingxiulong is known. There was also theropods like the smaller Panguraptor, and the large Sinosaurus which would have been a major pain for Xingxiulong. The potential Coelurosaur Eshanosaurus was present too, but this is doubtful and honestly it probably was just another prosauropod. Also present was Lukousaurus, which more likely than not is just a crocodilian. No ornithischians are known from the same environment as Xingxiulong, unfortunately.
Other: Xingxiulong, though very sauropod-like, was actually only midway on the sauropod family tree! Many animals, even in its own environment, were just as close to sauropods as Xingxiulong was. Still, Xingxiulong gives us a decent picture of how sauropods evolved at the Triassic-Jurassic transition.
~ By Meig Dickson
Sources under the Cut
Apaldetti, C.; Martinez, R.N.; Pol, D.; Souter, T. (2014). "Redescription of the Skull of Coloradisaurus brevis (Dinosauria, Sauropodomorpha) from the Late Triassic Los Colorados Formation of the Ischigualasto-Villa Union Basin, northwestern Argentina". Journal of Vertebrate Paleontology. 34 (5): 1113–1132.
Barrett, P.M.; Upchurch, P.; Wang, X.-L. (2005). "Cranial Osteology of Lufengosaurus huenei Young (Dinosauria: Prosauropoda) from the Lower Jurassic of Yunnan, People's Republic of China". Journal of Vertebrate Paleontology. 25 (4): 806–822.
Barrett, P.M.; Upchurch, P.; Zhou, X.-D.; Wang, X.-L. (2007). "The skull of Yunnanosaurus huangi Young, 1942 (Dinosauria: Prosauropoda) from the Lower Lufeng Formation (Lower Jurassic) of Yunnan, China". Zoological Journal of the Linnean Society. 150 (2): 319–341.
Lu, J.; Kobayashi, Y.; Li, T.; Zhong, S. (2010). "A New Basal Sauropod Dinosaur from the Lufeng Basin, Yunnan Province, Southwestern China". Acta Geologica Sinica. 84 (6): 1336–1342.
Luo, Z.; Wu, X.-C. (1997). "The small tetrapods of the Lower Lufeng Formation, Yunnan, China". In Fraser, N.C.; Sues, H.-D. (eds.). In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods. Cambridge: Cambridge University Press. pp. 251–270.
Martinez, R.N. (2009). "Adeopapposaurus mognai, gen. et sp. nov. (Dinosauria: Sauropodomorpha), with comments on adaptations of basal Sauropodomorpha". Journal of Vertebrate Paleontology. 29 (1): 142–164.
McPhee, B.W.; Yates, A.M.; Choiniere, J.N.; Abdala, F. (2014). "The complete anatomy and phylogenetic relationships of Antetonitrus ingenipes (Sauropodiformes, Dinosauria): implications for the origins of Sauropoda". Zoological Journal of the Linnean Society. 171 (1): 151–205.
McPhee, B.W.; Bonnan, M.F.; Yates, A.M.; Neveling, J.; Choiniere, J.N. (2015). "A new basal sauropod from the pre-Toarcian Jurassic of South Africa: evidence of niche-partitioning at the sauropodomorph–sauropod boundary?". Scientific Reports. 5: 13224.
McPhee, B.W.; Choiniere, J.N.; Yates, A.M.; Viglietti, P.A. (2015). "A second species of Eucnemesaurus Van Hoepen, 1920 (Dinosauria, Sauropodomorpha): new information on the diversity and evolution of the sauropodomorph fauna of South Africa's lower Elliot Formation (latest Triassic)". Journal of Vertebrate Paleontology. 35 (5): e980504.
Otero, A.; Pol, D. (2013). "Postcranial Anatomy and Phylogenetic Relationships of Mussaurus patagonicus (Dinosauria, Sauropodomorpha)". Journal of Vertebrate Paleontology. 33 (5): 1138–1168.
Pol, D.; Garrido, A.; Cerda, I.A. (2011). "A New Sauropodomorph Dinosaur from the Early Jurassic of Patagonia and the Origin and Evolution of the Sauropod-type Sacrum". PLoS ONE. 6 (1): e14572.
Prieto-Marquez, A.; Norell, M. (2011). "Redescription of a nearly complete skull of Plateosaurus (Dinosauria, Sauropodomorpha) from the late Triassic of Trossingen (Germany)". American Museum Novitates. 3727: 1–58.
Sues, H.-D.; Reisz, R.R.; Hinic, S.; Raath, M.A. (2004). "On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens formations (Lower Jurassic) of South Africa". Annals of Carnegie Museum. 73 (4): 239–257.
Wang, Y.-M.; You, H.-L.; Wang, T. (2017). "A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China". Scientific Reports. 73: 41881.
Wu, X.-C.; Currie, P.J.; Dong, Z.; Pan, S.; Wang, T. (2009). "A New Theropod Dinosaur from the Middle Jurassic of Lufeng, Yunnan, China". Acta Geologica Sinica. 83 (1): 9–24.
Xing, L.; Rothschild, B.M.; Ran, H.; Miyashita, T.; Persons, W.S.; Sekiya, T.; Zhang, J.; Wang, T.; Dong, Z. (2015). "Vertebral fusion in two Early Jurassic sauropodomorph dinosaurs from the Lufeng Formation of Yunnan, China". Acta Palaeontologica Polonica. 60 (3): 643–649.
Xu, X.; Zhao, X.; Clark, J.M. (2001). "A New Therizinosaur from the Lower Jurassic Lower Lufeng Formation of Yunnan, China". Journal of Vertebrate Paleontology. 21 (3): 477–483.
Yates, A.M.; Bonnan, M.F.; Neveling, J.; Chinsamy, A.; Blackbeard, M.G. (2009). "A new transitional sauropodomorph dinosaur from the Early Jurassic of South Africa and the evolution of sauropod feeding and quadrupedalism". Proceedings of the Royal Society B: Biological Sciences. 277 (1682): 787–794.
#Xingxiulong chengi#Xingxiulong#sauropodomorph#prosauropod#Dinosaur#Dinosaurs#Terrestrial Tuesday#Herbivore#Eurasia#Jurassic#Prehistoric Life#Prehistory#Paleontology#Palaeoblr#Factfile#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
208 notes
·
View notes
Text
Notocolossus gonzalezparejasi

By Ripley Cook
Etymology: Southern Giant
First described By: González Riga et al., 2018
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Macronaria, Titanosauriformes, Somphospondyli, Titanosauria, Lithostrotia
Status: Extinct
Time and Place: Between 88 and 86 million years ago, in the Coniacian of the Late Cretaceous


Notocolossus is known from the Plottier Formation of Argentina
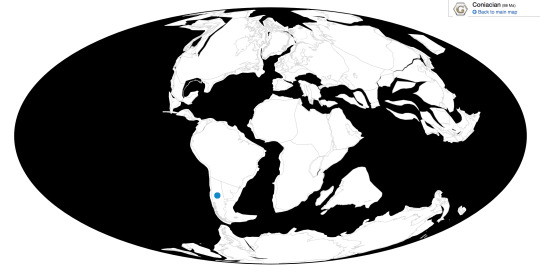
Physical Description: Notocolossus was a giant titanosaur, the group of dinosaurs that contains the largest land animals and most of the sauropods of the Cretaceous period. Like other titanosaurs, Notocolossus had a long thin neck, shorter (but still thin) tail, and extremely round stomach. I tend to describe titanosaurs to people on my tours as Bowling Balls with a noodle on either end, and Notocolossus would have followed that pattern. It probably was more than 25 meters in length, and it weighed as much as 60.4 tonnes. It had pillar-like feet - the front limbs had no toes whatsoever, essentially rendering them into columns; the back feet were reducing their toes as well, turning them into more column-like than ancestral sauropod feet. It had strong legs in addition to the sturdy feet, to help them stay upright with their extensive weight. The vertebrae were well fortified, to help to keep the animal supported. It would have been entirely scaly, and possibly had interesting color patterns for display to other members of the herd.
By González et al., CC BY 4.0
Diet: As a large titanosaur, Notocolossus would have primarily fed upon high levels of vegetation, especially upper levels of trees.
Behavior: Notocolossus appears to have been very common in its ecosystem, and thus it was probably a herding animal. These herds would spend most of their time scouring the open landscape for food, searching for plants and trees to feed on. They’d strip the plants of their leaves, and let them digest and break down in their stomachs over long periods of time. Being so large, Notocolossus probably didn’t have to defend itself from predators much; younger members of the herd and older members would have been vulnerable, but in general they were too big to be preyed upon. They were fairly active, being powered by a weird mixture of gigantothermy (body temperature maintained through size) and endothermy (body temperature maintained through one’s own energy); however, it seems unlikely that they took care of their young, given their body size and the nesting mod of other titanosaurs. That doesn’t mean it is certain either way, but it seems likely that they laid the eggs, and the surviving young only joined when they were big enough to keep up with the herd.

By Nobu Tamura, CC BY-SA 4.0
Ecosystem: The Plottier formation was an extremely large, semi-arid floodplain, with meandering rivers flowing through the area. These rivers sometimes swelled up, but were often very meager indeed, especially during the dry seasons. This lead to the ecosystem being filled with drier sediment - sand and red dirt - rather than mud. Still, plantlife was present here, including mosses, ferns, cycads, and lichens. It is even possible that some flowering plants were present. This environment was fairly lousy with titanosaurs - Petrobrasaurus, Muyelensaurus, and “Antarctosaurus” were all present, though Notocolossus was the most common of them. So, clearly, there were enough plants in the area for all of these animals to find food. Crocodylomorphs, predatory dinosaurs, and potential fish-eating raptors were present too, though none have been named. Many turtles also lived in this environment.
Other: Notocolossus is especially important because it actually preserved the foot bones, which is not common for titanosaurs; these foot bones show that the hindlimbs were especially adapted for holding up the large weight of the titanosaurs. The hands reduced their fingers and essentially became pillars, while the feet (hind limbs) also reduced the toe claws and compacted the toes themselves for better ability to support the weight. Notocolossus appears to have been closely related to other giant titanosaurs such as Patagotitan and Puertasaurus.
~ By Meig Dickson
Sources under the Cut
Carballido, José L.; et al. (2017). "A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs". Proc. R. Soc. B. 284 (1860): 20171219.
Chiappe, Luis M. (1993): Enantiornithine (Aves) Tarsometatarsi from the Cretaceous Lecho Formation of Northwestern Argentina. American Museum Novitates 3083: 1-27. [English with Spanish abstract].
Coria, Rodolfo A.; Currie, Philip J.; Eberth, David & Garrido, Alberto (2002): Bird footprints from the Anacleto Formation (Late Cretaceous), Neuquén, Argentina. Ameghiniana 39(4): 453-463.
Fossa Mancini, E.; E. Feruglio, and J.C. Yussen de Campana. 1938. Una reunión de geólogos de YPF y el problema de la terminología estratigráfica ("A YPF geologists' reunion and the problem of stratigraphy terminology"). Boletín de Informaciones Petroleras 15. 1–67.
González Riga, Bernardo J.; Lamanna, Matthew C.; Ortiz David, Leonardo D.; Calvo, Jorge O.; Coria, Juan P. (2016). "A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot". Scientific Reports. 6: 19165.
Gonzalez Riga, B.J.; Mannion, P.D.; Poropat, S.F.; Ortiz David, L.; Coria, J.P. (2018). "Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships" (PDF). Journal of the Linnean Society. 184 (1): 136–181.
Leanza, Héctor A.; et al. (2004). "Cretaceous terrestrial beds from the Neuquén Basin (Argentina) and their tetrapod assemblages". Cretaceous Research. 25 (1): 61–87.
Lockley, Martin; Matsukawa, Masaki; Ohira, Hiroto; Li, Jianjun; Wright, Joanna; White, Diane & Chen, Peiji (2006): Bird tracks from Liaoning Province, China: New insights into avian evolution during the Jurassic-Cretaceous transition. Cretaceous Research 27(1): 33-43.
Musacchio, E. A., P. Vallati. 2007. LATE CRETACEOUS NON MARINE MICROFOSSILS OF THE PLOTTIER FORMATION (CRETACEOUS) AT ZAMPAL, ARGENTINA. 4th European Meeting on the Palaeontology and Stratigraphy of Latin America: 273 - 278.
Salgado, L.; Coria, R.A. & Chiappe, Luis M. (2005): Osteology of the sauropod embryos from the Upper Cretaceous of Patagonia. Acta Palaeontologica Polonica 50(1): 79–92.
Sánchez, María Lidia; Susana Heredia, and Jorge O. Calvo. 2006. Paleoambientes sedimentarios del Cretácico Superior de la Formación Plottier (Grupo Neuquén), Departamento Confluencia, Neuquén (Sedimentary paleoenvironments in the Upper Cretaceous Plottier Formation (Neuquen Group), Confluencia, Neuquén). Revista de la Asociación Geológica Argentina 61. 3–18.
#Notocolossus gonzalezparejasi#Notocolossus#dinosaur#titanosaur#prehistoric life#paleontology#prehistory#palaeoblr#factfile#Mesozoic Monday#herbivore#south america#cretaceous#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
89 notes
·
View notes
Text
Amygdalodon patagonicus

By José Carlos Cortés
Etymology: Almond Tooth
First Described By: Cabrera, 1947
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda
Status: Extinct
Time and Place: Between 180 and 175 million years ago, in the Toarcian of the Early Jurassic

Amygdalodon is known from the Cerro Carnerero Formation of Chubut, Argentina

Physical Description: Amygdalodon is a fairly poorly known early Sauropod, known really only from bone fragments and teeth. As an early sauropod, though, it would have been somewhat bulky, and probably somewhere around 12 meters long. It would have walked around on all fours, and very similar to other early branching sauropods. It would have had a semi long neck and long tail, but no extreme specializations either way. It would have probably been entirely scaly due to its large body size.
Diet: Amygdalodon would have been primarily herbivorous.
Behavior: It is uncertain what the behavior of Amygdalodon would have been, given we don’t have a lot of fossil evidence of this dinosaur. That being said, it probably spent most of its day feeding, and most likely lived in large groups of other Amygdalodon for protection. It probably also took care of its young, and had an active metabolism - but, again, this is just conjecture.

By Ripley Cook
Ecosystem: Amygdalodon is known from a coastal environment, a pebbly beach associated with sand as the pebbles were eroded from the waves of the sea; Amygdalodon itself mainly keeping to the rivers as they emptied out into the ocean. Unfortunately, no other dinosaurs were found alongside Amygdalodon; only one poorly preserved shrimp. So it is uncertain what sorts of plants Amygdalodon lived alongside, what other animals it interacted with, and what may have preyed upon it.
Other: Amygdalodon is the most basal known sauropod from the continent of South America!
~ By Meig Dickson
Sources under the Cut
Cabrera, A. 1947. Un saurópodo nuevo del Jurásico de Patagonia. Instituto del Museo de la Universidad Nacional de La Plata, Notas del Mueso de La Plata, Paleontología 12(95):1-17.
Carballido, J. L., and D. Pol. 2010. The dentition of Amygdalodon patagonicus (Dinosauria: Sauropoda) and the dental evolution in basal sauropods. Comptes Rendus Palevol 9:83-93
Cortés, José María. 1986. Estratigrafía y tectónica de la región central de la Provincia de Chubut, República Argentina (PhD thesis), 1–242. Universidad de Buenos Aires.
Holwerda, F. M., & D. Pol (2018). Phylogenetic analysis of Gondwanan basal eusauropods from the Early-Middle Jurassic of Patagonia, Argentina. Spanish Journal of Palaeontology 33(2): 289-298.
Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 176
Rauhut OWM. Revision of Amygdalodon patagonicus Cabrera, 1947 (Dinosauria, Sauropoda). Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe. 2003;6:173–181.
#Amygdalodon#Amygdalodon patagonicus#Sauropod#Dinosaur#Prehistoric Life#Palaeontology#Prehistory#Palaeoblr#Dinosaurs#Factfile#Jurassic#South America#Herbivore#Mesozoic Monday#paleontology#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
150 notes
·
View notes
Text
Malawisaurus dixeyi

By Ripley Cook
Etymology: Reptile from Malawi
First Described By: Jacobs et al., 1993
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Macronaria, Titanosauriformes, Somphospondyli, Titanosauria, Lithostrotia
Status: Extinct
Time and Place: Sometime between 125 and 113 million years ago, in the Aptian age of the Early Cretaceous

Malawisaurus is known from the Upper Member of the Dinosaur Beds Formation in Mwakasyunguti, Malawi
Physical Description: Malawisaurus was a titanosaur - the kind of heavy-set, bulky sauropods that dominated the Cretaceous landscape. Though it did not get as big as its relatives, it is among the group of the largest land animals to walk the earth. Still, Malawisaurus was on the smaller size, probably only reaching 4.7 tons, and maybe around 16 meters long. Malawisaurus had a long neck and tail, and a distinctively short, squat, and squarish head - with very large nasal holes in the skull. This would have helped to keep its skull relatively cool, an important feature for a large animal in a hot and humid climate.

By Slate Weasel, in the Public Domain
Despite having fossil material of the skull and teeth - a rarity for titanosaurs - redescriptions and reexaminations of Malawisaurus have shown that the wealth of material assigned to it might not all be one species, so studies to clear up the picture are ongoing. For now, it seems as though there were two distinct morphs of Malawisaurus, differing based on variations in the vertebrae of the neck. The tail vertebrae also had very long connections between one another, that would have made its tail a little more stiff - possibly for balance. There were also depressions in some of the sides of the vertebrae, though it’s uncertain what they would have been for.
The braincase of Malawisaurus has been thoroughly studied, helping us to understand more about its brain and, thus, behavior. Malawisaurus had a very large semicircular canal, like other early titanosaurs. It also had a nerve for the eye much more to the side than seen in other sauropods. Malawisaurus had thick, peg-like teeth, which would have aided in stripping leaves from branches.

By Mesozoical, CC BY-SA 3.0
As a Titanosaur, it would have been largely scaly; it possible it may have had small scutes covering the skin to aid in protecting its body from predators.
Diet: Malawisaurus would have been a medium-level browser of plant material.
Behavior: Based on the head of Malawisaurus, it would have probably moved its neck and head slowly from side to side, rather than quickly; this would have made it very sluggish as it moved its head along the plant level to browse, though this probably would have worked out, as it had to eat a lot of plant material every day to keep its size up. This means it would have browsed, slowly and surely, through the row of leaves, before moving on to the next one. Malawisaurus would have probably maintained a fairly active metabolism through sheer body size, and would have needed to eat a lot of plant material to uphold itself each day. It would have moved its eyes only a little to see more of the area around it from side to side, similar to its relative Sarmientosaurus. This means that it also would have moved its eyes slowly, in addition to its head and neck.

By José Carlos Cortés
Malawisaurus probably wouldn’t have been the fastest of dinosaurs in general, though of course it’s difficult to tell for certain. It was probably fairly common, too, and may have lived in herds. Whether or not it would have been involved in any parental care is uncertain, though it probably would have laid its eggs in mounds like other titanosaurs.
Ecosystem: Malawisaurus is known from the Dinosaur Beds Environment, a habitat of a river bed flowing from a series of lakes. This was an iron-rich muddy environment near the rivers that transitioned to a more sandy habitat near the lakes, with the entire ecosystem being in a somewhat more temperate climate - being fairly far from the equator. Still, a lack of major palynological studies leaves us wondering what sorts of plants were in the area. Here there were a wide variety of crustaceans, frogs, and ray-finned fish, as well as the turtle Platycheloides, the notosuchian Malawisuchus, and another titanosaur, Karongasaurus. There seems to have been a third type of dinosaur, possibly another kind of Malawisaurus as per the two morphs - indicating the presence of a diverse, but endemic, clade of sauropods in this environment. So, clearly, there was enough plant material to feed at least three large dinosaurs; hopefully one day we’ll know exactly what.

By Scott Reid
Other: The braincase of Malawisaurus indicates it was a basal titanosaur, being closer to the base of the evolutionary tree for this group. It is one of the only titanosaurs for whom a skull is known; which makes the skeleton being kind of useless all the sadder.
~ By Meig Dickson
Sources under the Cut
Andrezejewski, K., M. J. Polcyn, D. A. Winkler, E. G. Chindebvu, L. L. Jacobs. 2019. The braincase of Malawisaurus dixeyi (Sauropoda: Titanosauria): A 3D Reconstruction of the brain endocast and inner ear. PLoS ONE 14 (2): e0211423.
Dixey, F. 1925. The discovery of fossil reptiles. Annual Report of the Geological Survey Department for the Year 1924, Nyasaland Protectorate 1924:7
Gomani, E. M. 1999. Sauropod caudal vertebrae from Malawi, Africa. In Y. Tomida, T. H. Rich, and P. Vickers-Rich (eds.), Proceedings of the Second Gondwanan Dinosaur Symposium, National Science Museum Monographs 15:235-248
Gorscak, E. 2016. Descriptive and Comparative Morphology of African Titanosaurian Sauropods: New Information on the Evolution of Cretaceous African Continental Faunas. A dissertation presented to the faculty of the College of Arts and Sciences of Ohio University.
Filippi, L. S., L. Salgado, A. C. Garrido. 2019. A new giant basal titanosaur sauropod in the Upper Cretaceous (Coniacian) of the Neuquén Basin, Argentina. Cretaceous Research 100: 61 - 81.
Haughton, S. H. 1928. On some reptilian remains from the Dinosaur Beds of Nyasaland. Transactions of the Royal Society of South Africa 16:67-75
Jacobs, L. L., D. A. Winkler, Z.M. Kaufulu and W.R. Downs. 1990. The Dinosaur Beds of northern Malawi, Africa. National Geographic Research 6(2):196-204
Jacobs, L. L., D. A. Winkler, W. R. Downs and E. M. Gomani. 1993. New material of an Early Cretaceous titanosaurid sauropod dinosaur from Malawi. Palaeontology 36(3):523-534 \
Paul, Gregory S. (2010) The Princeton Field Guide to Dinosaurs.
Tidwell, V., Carpenter, K. & Meyer, S. 2001. New Titanosauriform (Sauropoda) from the Poison Strip Member of the Cedar Mountain Formation (Lower Cretaceous), Utah. In: Mesozoic Vertebrate Life. D. H. Tanke & K. Carpenter (eds.). Indiana University Press, Eds. D.H. Tanke & K. Carpenter. Indiana University Press. 139-165.
#Malawisaurus#Malawisaurus dixeyi#titanosaur#sauropod#dinosaur#palaeoblr#factfile#paleontology#prehistoric life#prehistory#Cretaceous#Africa#Herbivore#Mesozoic Monday#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
158 notes
·
View notes
Text
Normanniasaurus genceyi

By Scott Reid
Etymology: Reptile from Normannia
First Described By: Le Loeuff et al., 2013
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Macronaria, Titanosauriformes, Somphospondyli, Titanosauria
Status: Extinct
Time and Place: Between 113 and 107 million years ago, in the Albian age of the Early Cretaceous

Normanniasaurus is known from the Poudingue Ferrugineux Formation of northwestern France

Physical Description: Normanniasaurus was a fairly early titanosaur, the group of Cretaceous period sauropods that make up the greatest diversity of the group. Normanniasaurus itself was a small to medium sized sauropod, probably around 12 meters long. It was interesting in a few ways - notably, it had the ball-and-socket connections between vertebrae, like later titanosaurs, but only at the base of the tail. This would make the tail fairly flexible compared to non-titanosaur sauropods, but not as flexible as later forms. Unlike later titanosaurs, however, it had a very stiff back - more like more basal, non-titanosaur sauropods. The spines of Normanniasaurus also had huge connected holes on the tail. In all, Normanniasaurus showcases a mixture of basal and derived sauropod traits, indicative of its early position in titanosaur evolution.
Diet: As a smaller titanosaur, Normanniasaurus was probably a mid-level browser, though it’s possible it could reach higher levels of vegetation when needed.

By José Carlos Cortés
Behavior: Being still small-ish in size, Normanniasaurus would have probably been more skittish than larger titanosaurs, needing to react quickly in the presence of potential danger, as it couldn’t rely on large size for it (given the presence of larger predators in its area, which is uncertain). It spent most of its day grazing on food, storing it all in a large stomach. As only a few fossils of Normanniasaurus have been found, it’s difficult to tell if it lived in herds, but it seems somewhat likely given its smaller size and need to protect itself. It is unknown whether or not it would have cared for its young.
Ecosystem: The Poudingue Ferrugineux Formation formed along a coastal environment, meaning Normanniasaurus would have interacted with the ocean ecosystem moreso than more inland animals. However, no other dinosaurs have been found from this formation, much less other fossils, so it’s difficult to tell what would have been the predators of Normanniasaurus specifically, or competitors, or just what it would have interacted with at all. There do seem to be other titanosaurs from the general time and place, including Aepisaurus and Macrurosaurus, implying that Normanniasaurus might have been one member of a fairly diverse sauropod-heavy ecosystem.
Other: Normanniasaurus is one of the earliest known titanosaurs, though it’s precise placement in the group is murky. It might be the oldest known Aelosaur, which would also make it the only member of that group known from Europe. Alternatively, it might be more like Malawisaurus, which lived at the same time, but in Africa. If its a basal titanosaur, it’s fairly typical for the group; but if it is an Aeolosaur, it might have lost some titanosaur traits, which would make it a fascinating case in titanosaur evolution.
~ By Meig Dickson
Sources under the Cut
Buffetaut, E. 1984. Une vertèbre de dinosaurien sauropode dans le Crétacé du Cap de la Hève (Normandie). Actes du Muséum d’Histoire naturelle de Rouen 7: 215 - 221.
Le Loeuff, J., S. Suteethorn, E. Buffetaut. 2013. A new sauropod dinosaur from the Albian of Le Havre (Normandy, France). Oryctos 10: 23 - 30.
#Normanniasaurus#Titanosaur#Dinosaur#Palaeoblr#Normanniasaurus genceyi#factfile#Cretaceous#Terrestrial Tuesday#Herbivore#Eurasia#paleontology#prehistory#prehistoric life#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
254 notes
·
View notes
Text
Qingxiusaurus youjiangensis

By Scott Reid
Etymology: Reptile from Qinxiu
First Described By: Mo et al., 2008
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Macronaria, Titanosauriformes, Somphospondyli, Titanosauria
Status: Extinct
Time and Place: Sometime between 100 and 66 million years ago, in the Late Cretaceous period (exact timing is uncertain)

Qingxiusaurus is known from the Dashi Site in Guanxi, China
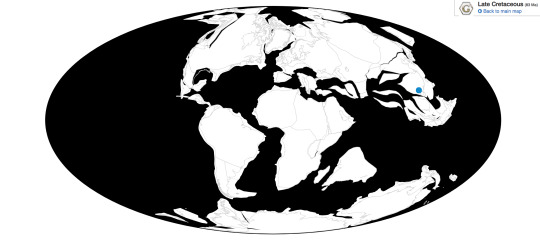
Physical Description: Qingxiusaurus is a titanosaur, a type of heavy-set derived sauropod that dominated the landscape of the Cretaceous Period. Thus, we can assume that Qingxiusaurus was similar to other titanosaurs - large, wide-set, and with legs like pillars for holding up its bulk. Beyond that, however, it is a mystery. It is known from only parts of the legs and neural spines, which don’t say much about it; it does have flat, paddle-shaped spines, which is a bit weird, but it is uncertain what that would mean in terms of lifestyle.
Diet: Qingxiusaurus would have been either a mid-level or upper-level browser of plant material.
Behavior: Behavior of Qingxiusaurus is a mystery, as we know little about its size, shape, sociality, or environment.
Ecosystem: Little is known of where Qingxiusaurus lived, but it seems to have been a fairly muddy environment, perhaps an often-flooded floodplain or a swamp. Qingxiusaurus lived with Naningosaurus, an early hadrosaur; the predators of the environment are not known.
Other: Qingxiusaurus shares many features with Saltosaurids, but it’s uncertain if it’s actually a member of that group.
~ By Meig Dickson
Sources under the Cut
D'Emic, M. D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society 166:624-671
Mannion, P. D., and J. O. Calvo. 2011. Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian–early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: implications for titanosaur systematics. Zoological Journal of the Linnean Society 163:155-181
Mo, J.-Y., C.-L. Huang, Z.-R. Zhao, W. Wang, and X. Xu. 2008. A new titanosaur (Dinosauria: Sauropoda) from the Late Cretaceous of Guangxi, China. Vertebrata PalAsiatica 46(2):147-156
Upchurch, Paul; Barrett, Paul M.; Dodson, Peter. (2004). "Sauropoda". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 259–322.
Wilson, J. A., and P. Upchurch. 2009. Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Early Cretaceous of China. Journal of Systematic Palaeontology 7(2):199-239
#Qingxiusaurus#Qingxiusaurus youjiangensis#Titanosaur#Dinosaur#Paleoblr#Factfile#Dinosaurs#Cretaceous#Eurasia#Herbivore#Mesozoic Monday#Prehistoric Life#Prehistory#Paleontology#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
82 notes
·
View notes
Text
Shingopana songwensis

By José Carlos Cortés
Etymology: Wide Neck
First Described By: Gorscak et al., 2017
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Macronaria, Titanosauriformes, Somphospondyli, Titanosauria, Lithostrotia, Aeolosaurini
Status: Extinct
Time and Place: Between 83.6 and 72.1 million years ago, in the Campanian age of the Early Cretaceous
Shingopana is known from the Namba Member of the Galula Formation in southwestern Tanzania

Physical Description: Shingopana is a small titanosaur - the group of long-necked dinosaurs that lived the longest (through the end of the Cretaceous period) and grew to be the largest. Shingopana, however, showcases the group’s diversity - they did a lot of weird things, not all of them enormous size. Shingopana was about 8 meters long, and only about two meters tall at the hip - when you compare that to the largest titanosaurs, which could reach up to 30 meters in length, this is quite small indeed. Shingopana is known from some vertebrae, a limb bone, and some ribs. The vertebrae show a weird trait found in many titanosaurs - rather than having, well, regular ol’ spines on the backs of the vertebrae, Shingopana had instead huge balls of rough bone. This probably helped Shingopana strengthen its neck, giving the muscles more attachment sites. The ribs have weird flanging edges, though the function of this is not entirely clear. As for the legs, it seems that they had fewer attachment sites for muscles, making them less beefy than your average titanosaur’s limbs. Unfortunately, more specific proportions such as tail and neck length and limb length is not known without further material from this titanosaur. Like other titanosaurs, it would have had a large stomach; and it probably had two rows of round osteoderms running down its back.
Diet: As a titanosaur, Shingopana would have been entirely herbivorous, feeding on a relatively large volume of plant material every day. The smaller size of Shingopana would indicate it was a medium-level browser, eating mainly cycads and conifer trees, but not high up in the tree level.
Behavior: Shingopana, as a fairly small titanosaur, would have been a moderately skittish sort of animal, as it wouldn’t have been able to defend itself with size from predators as well as bigger sauropods. It would have spent most of its time during the day grazing on food, storing it all in a very large stomach. It is uncertain whether or not Shingopana would have lived in herds, but it seems at least somewhat likely that it would have traveled in groups. It is unknown whether or not it would have cared for its young.

By Scott Reid
Ecosystem: Shingopana is known from the Namba Member of the Galula Formation, and it wasn’t the only titanosaur in the environment - it lived alongside the larger Rukwatitan, indicating niche partitioning between the two (the niches being… high plants and low plants). There was also the dog-like crocodyliform Rukwasuchus, that probably would have fed upon Shingopana in some capacity, as it was a medium-sized land-based carnivore. There was also the weirdly mammal like Pakasuchus, another crocodyliform with a dog-like nose and teeth - even though it wasn’t closely related to Rukwasuchus, the two looked very similar indeed. However, Pakasuchus was a little smaller than Rukwasuchus, and would have mainly been a threat to young Shingopana. Beyond that, there are multiple fish, theropods, and even potential close-to-birds, but they haven’t been named. This environment was a river system, though the plant life of the area is not well known.
Other: Shingopana is notable for being an Aeolosaurin - a group of titanosaurs that are only known from South America, except for Shingopana, which is known from Africa. This indicates that the range of this group is larger than previously known, though it is uncertain how widespread they were in Africa without more fossils found. Aeolosaurins are notable for having weirdly shaped tails, though without fossils of the tail vertebrae it is uncertain if Shingopana also had a weird tail.
~ By Meig Dickson
Sources under the cut
Gorscak, E., P. M. O’Connor, E. M. Roberts, N. J. Stevens. 2017. The second titanosaurian (Dinosauria: Sauropoda) from the middle Cretaceous Galula Formation, southwestern Tanzania, with remarks on African titanosaurian diversity. Journal of Vertebrate Paleontology: 21343250.
Gorscak, E., P. M. O’Connor. 2019. A new African Titanosaurian Sauropod Dinosaur from the middle Cretaceous Galula Formation (Mtuka Member), Rukwa Rift Basin, Southwestern Tanzania. PLoS ONE 14 (2): e0211412.
Mocho, P., A. Perez-García, M. Martín Jimenez, F. Ortega. 2019. New remains from the Spanish Cenomanian shed light on the Gondwanan origin of European Early Cretaceous titanosaurs. Cretaceous Research 95: 164 - 190.
Prasad, V., C. A. E. Strömberg, H. Alimohammadian, A. Sahni. 2005. Dinosaur Coprolites and the Early Evolution of Grasses and Grazers. Science 310 (5751): 1177 - 1180.
Sertich, J. J. W., P. M. O'Connor. 2014. A new crocodyliform from the middle Cretaceous Galula Formation, southwestern Tanzania. 34(3):576-596
Widlansky, S. J., W. C. Clyde, P. M. O’Connor, E. M. Roberts, N. J. Stevens. 2017. Paleomagnetism of the Cretaceous Galula formation and implications for vertebrate evolution. Journal of African Earth Sciences: 10.1016/j.jafrearsci.2017.11.029.
#shingopana#shingopana songwensis#dinosaur#palaeoblr#titanosaur#factfile#dinosaurs#prehistoric life#prehistory#paleontology#cretaceous#africa#herbivore#mesozoic monday#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
117 notes
·
View notes
Text
“Microdontosaurus”
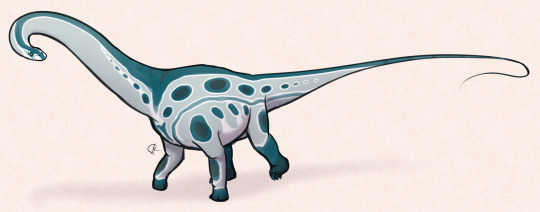
By José Carlos Cortés
Etymology: Small Tooth Reptile
First Mentioned By: Zhao, 1983
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Plateosauromorpha, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda
Status: Extinct
Time and Place: “Microdontosaurus” lived sometime in the Middle Jurassic epoch, between 174.1 and 163.5 million years ago, though it is uncertain exactly when.
“Microdontosaurus” is known from the Dabuka Formation of Tibet, which is currently a part of China.
Physical Description: “Microdontosaurus” is known from a specimen that hasn’t yet been described, but is only stated as a sauropod of some sort. It had small teeth, which presumably indicates that it would have used these teeth in stripping off finer plant material. Given that it hasn’t really been classified in any way, the only thing that can be said for its general appearance is that, as a sauropod, it would have been a quadrupedal animal, with a long tail and long neck, as well as a small head.
Diet: Leaves and other plant material; potentially finer leaves and shoots given its supposed small teeth size. It is uncertain what level of browser “Microdontosaurus” was.
Behavior: Unknown
Ecosystem: The ecology of the Dabuka Formation is currently unknown; though, as it was the Jurassic, it probably would have been a tropical and wet environment. Other dinosaurs present in the area included a theropod closely related to Megalosaurus, the also unofficially named “Ngexisaurus” theropod that was probably similar to Ceratosaurus, another informally described Sauropod “Lancanjiangosaurus”, and the informally named stegosaur “Changdusaurus”. This Tibetan formation is just, filled, with not properly described dinosaurs.
Other: As a nomen nudum, “Microdontosaurus” has not been officially named; as such, little can be said about it scientifically speaking. It may be closely related to animals such as Brachiosaurus, but that’s just conjecture. It also may have been named as a different dinosaur, though of course, no one is sure.
~ By Meig Dickson
Sources under the cut
Anonymous. 1977. Dinosaurs found in Tibet. New Scientist 73 (1038): 329.
Gilmore, 1902. Discovery of teeth in Baptanodon, an ichthyosaurian from the Jurassic of Wyoming. Science. 16(414), 913-914.
Zhao, 1985. The reptilian fauna of the Jurassic in China. Pgs. 286 - 289 in Wang, Cheng and Wang (eds.). The Jurassic System of China. Geological Publishing House, Beijing.
#microdontosaurus#dinosaur#sauropod#palaeoblr#herbivore#eurasia#mesozoic monday#jurassic#sauropodomorph#paleontology#prehistoric life#prehistory#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#factfile
123 notes
·
View notes
Text
Fulengia youngi

By Tas on @i-draws-dinosaurs
PLEASE SUPPORT US ON PATREON. EACH and EVERY DONATION helps to keep this blog running! Any amount, even ONE DOLLAR is APPRECIATED! IF YOU ENJOY THIS CONTENT, please CONSIDER DONATING!
Name: Fulengia youngi
Name Meaning: Anagram of Lufeng
First Described: 1977
Described By: Carroll and Galton
Classification: Dinosauria, Saurischia, Eusaurischia, Sauropodmorpha, Plateosauria, Massopoda, Massospondylidae
HAHAHAHAHHA GUESS WHAT WE’RE NEVER FREE FROM BITCHES THAT’S RIGHT IT’S THE PROSAUROPODS
Fulengia is probably Lufengosaurus. It is known from very fragmentary remains from the Lufeng Formation of Yunnan Province, China, living around 190 million years ago, in the Hettangian to Sinemurian ages of the Early Jurassic. For a while it was thought to be a lizard, but is now back to being a prosauropod, probably synonymous with Lufengosaurus, though we can’t know for sure, because the fossil is terrible.
Sources:
http://www.dinochecker.com/dinosaurs/FULENGIA
https://en.wikipedia.org/wiki/Fulengia
This is your friendly reminder that ADAD relies on reblogs up and until @staff gets it’s shit together. Please reblog this post.
Shout out goes to @weholddear!
#fulengia#fulengia youngi#dinosaur#prosauropod#palaeoblr#weholddear#paleontology#prehistory#prehistoric life#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#factfile#Dìneasar#דינוזאור#डायनासोर#ديناصور#ডাইনোসর#risaeðla#ڈایناسور#deinosor#恐龍#恐龙#динозавр#dinosaurio
53 notes
·
View notes
Last Seen Blogs




