#genotypic
Text
Ecological consequences of genotypic diversity within a foundation plant, Spartina alterniflora, are pervasive but not universal across multiple stress gradients
5 notes
·
View notes
Text
Jada Stevens In Twerk For Cum
Paige Turnah Shaking Her Thicc Booty
Indian bangali college girl sex with bf
Gay emo s porn video and nude young boy Is it possible to be in enjoy
Movie straight gay man seduced xxx Desperate man does anything for
One of my thots like creampie riding
Petite Latina gets picked up and fucked in Columbia
Indian adult web serial sex scenes
Young Teens House Party best with NO PANTIES
Interracial Fucking With Blonde Milf
#heart-shed#floodlet#nondesistence#hot-drawn#punnet#chipboard#concaver#genotypic#all-steel#aardwolves#lathesmen#blastopore#kirkify#outlinear#unitarism#nonconvergently#surfings#keelboatman#raylet#CLEF
0 notes
Text


btw protip: when making fankid designs, look at the concept art of the parents for inspiration
#you know not to blatant about this headcanon i like to think about for funsies but#naegiri#pretend that the concept designs are buried somewhere in their genotypes or somethin#more fun that way and you can get some interesting variations#danganronpa#rain code#makoto naegi#kyoko kirigiri#shuichi saihara#yuma kokohead#raincode
141 notes
·
View notes
Text
idk who needs this reminder but uh race is a social construct, ethnicity is not. we can use DNA to determine the ethnicity of someone post mortem, we cannot determine their race that way. using facial reconstruction techniques we can estimate what they may have looked like and compare that with definitions of race from their lifetime, but even that is shaky ground to walk on.
that being said: many ethnic groups do not rely on blood quantum and in fact many actively reject it (Indigenous peoples especially). ethnicity is not solely genetic. its culture, its language, its customs, its religion, its clothing. so when you see someone argue that the concept of a Jewish ethnicity is fake or propaganda, they are willfully ignoring the fact that Jewish culture in the diaspora is diverse and unique, and also makes diaspora Jews distinct from the populations of the countries they now live in. being Jewish significantly changes your experience of any given country because you blend your culture with your country’s. because of this, we have yiddish and ladino and other judeo-languages, we have diaspora groups, we have unique practices all over the world because of our unique ethnic background.
and still, if you sample DNA from a Jew in Romania and a Jew in England and a Jew in Ethiopia, their DNA will be considerably more similar than their own DNA compared to goyim from their home country. the ethno in ethnoreligion has meaning, and denying it minimizes Jewish identity at best.
#can u tell i have a degree in sociology#sociology of race and ethnicity#phenotype and genotype are different things!#blood quantum is racist bullshit!#but that doesn’t mean we should ignore the existence of genotypic ethnic markers!#they can be useful for anthropological reasons and also can influence a person’s life significantly#ex. i have a heart condition that is not only predominantly found in ashkenazi jews#but genome sequencing has allowed us to trace the gene mutation that caused this disease back to one (1) russian rabbi centuries ago#‘russian’ meaning what is now russia#idk what the political map looked like back then#anyway#sociological misinformation will not survive on my watch#jumblr
61 notes
·
View notes
Text


& who asked you two to stand so close???
#pressure test is always gonna be one of my fav thiam episodes cuz like#theo is being so protective but trying to hide it behind pissing liam off at the same time hes trying to be a realist to his idealist#and convince him to take care of himself first before other people 😭#me when i swear not to save my enemy but decide id rather face death for him multiple times than see him hurt by anyone including himself#IDIOT#theo positioning himself between monroe and liam when they leave the station . that marking the beginning of the 6b streak +#where they keep getting paired off together and learn how to work as a team#they were def attached at the hip the entire time the pack was in hiding the same way scalia and morey were#like… leaving the station. at the zoo. driving back to town together. theo following him to the school. waiting under that night to tell +#scott about what they found. and then they were both grumpy about getting split up during genotype like they havent been living in each +#others pockets for who knows how long atp …. they are so .#jeff davis you will pay for not showing me all of the bonding that happened during this time. I HATE YOU!#thiam#theo raeken#liam dunbar#theo raeken x liam dunbar#theo raeken/liam dunbar#theo x liam#theo/liam#6.15 pressure test#6.17 werewolves of london#teen wolf
121 notes
·
View notes
Note
do you use a font for your text? if so, what is it?
i use a lot of fonts! i'm something of a Fonts Connoisseur, heh....some of my beloved mutuals have often come to me asking for help identifying fonts cuz i have an uncanny talent for it *smiles*
BUT to actually answer your question:


#not art#you can probably fairly easily find all these through google or sites like dafont or the Y2K aesthetic institute#BTW the version of genotype i have [or maybe just the font period] doesn't have numbers so the 0046 in my new watermark is a different font#i forget which one exactly. a shocking amount of Y2K futurism style fonts just don't have numbers for some reason
23 notes
·
View notes
Text
Phenotype vs Genotype
Phenotype
-- the trait produced by the genotype
-- depends on both genotype and environmental factors
-- not inherited by offspring
-- consists of expressed genes
-- dominant alleles can mask recessive alleles
-- genotype AA causes phenotype A
-- genotype Aa causes phenptype A
-- genotype aa causes phenotype a
Genotype
-- the actual genetic code of an organism
-- codes for the phenotype
-- consists of both dominant and recessive alleles
-- inherited by offspring
-- usually refers to one specific gene causing a trait
-- written as a combination of uppercase and lowercase letters
-> AA
-> Bb
-> ee
-- dominant = uppercase letters
-- recessive = lowercase letters
-- both uppercase = homozygous dominant
-- one uppercase and one lowercase = heterozygous
-- both lowercase = homozygous recessive
.
Patreon
#studyblr#notes#biology#bio#biology notes#bio notes#life science#science#health science#genetics#genetics notes#genotype#phenotype#homozygosity#heterozygosity#genetic code#biological science#study guide#genetics study guide
21 notes
·
View notes
Text
A completely self-indulgent imaginary rework of the EMS system
The EMS is a good system. I like it. I'm glad it exists.
I still think it has stupid parts, and i'd change a few codes if I could.
1. Base colors
The letters are all over the place. Why is red and cream put between the eumelaninistic variant colors? Why is there a big gap in the lettering after j? I know historically why black is n, but i still don't like it. So my suggestion would be:
a - black
b - blue
c - chocolate
d - lilac
e - cinnamon
f - fawn
g - red
h - cream
w - white (white is a freak, it can get it's own letter from the end of the alphabet)
These are the ones usually deemed as base colors. If this ever changes, the new colors can get the letter i, j, k and l.
More simple genes get a color modifier letter after the base color's:
m - caramel
n - sunshine and other bimetal-type golden variants (corin)
o -
p - phoenix (if i'm right with it not changing the nose color)
q - karpati
r - recessive red variants (amber, russet, serdolic; i group them together for now, since they are all restricted to one breed and don't compete; if they become more widespread, they'd probably need an own number) (mc1r)
s - silver/smoke
t - tortoiseshell
Golden doesn't get a letter, because wide band is denoted in the numbering.
2. White spotting
This is the least organised part of the EMS coding. It'd completely overthrow it, and probably rework again when we finally have all the genetics understood.
01 - locket, belly spot ect (very low grade white, no ws allele)
02 - white attributed to DBE. ONLY if there's no other type of white spotting.
03 - glove (wgwg ONLY)
04 - low white (~09 now, and mitted and snowshoe are here too, doesn't get different code until proven different allele)
05 - intermediate white, bicolor
06 - high white, harlequin, van
09 - indeterminable white spotting
3. Wide band
11 - low wide band (golden/silver tabby)
12 - medium wide band (golden/silver shaded/tipped)
13 - high wide band (golden/silver shell/chinchilla)
A black golden ticked tabby would be a 11 25.
4. Tabby pattern
You know, i'm content with this one. I wouldn't touch it, it's good as it is.
5. Color restriction
Same. Mocha could be added as 34, but that'll likely happen anyway.
6. Charcoal
I'd introduce this as an extra letter. It'll probably need some rework when the genetics clear out.
41 - midnight charcoal
42 - twilight charcoal
So. For example a light amber tortoiseshell smoke bicolor would be brst 05, a cinnamon golden tipped tabby would be e 12, a black mink rosetted charcoal would be a 24 32 41.
#i know nothing like this will happen#the current EMS system with a few modifications will probably stay in use for until we just genotype all the pedigree cats at birth#(oh i do hope that'll happen and i'll live to see it)#but i guess i wanted to express my thought
9 notes
·
View notes
Text
no I will not put the matress away the matress is under the table and I am under the table and I am drawing my ocs making out while simoutaneously revising genetics. is there a problem. I am not leaving my under the table spot
#anyway does anyone wanna see my ocs making out. or the creepy marker woman I drew#or do you wanna learn about the genotypic ratio of a dihybrid cross#bcs I am up for showing you all of that 👍#lamp.txt
12 notes
·
View notes
Text
i thought too much about sex determination in dragons and now it's one am and i'm manic
color determination must be non-chromosomal, right, otherwise there's no genetic way *at all* to get all the spread of colors in the clutch... but then why is the ratio of F to M so perfectly 50-50... that's sooo easy to explain with XY or ZW or EVEN X0 system
but then why would there be two morphs of female with such ridiculously different hatch rates. i am having SO much trouble finding examples of species with female morphs, it seems like everything that does morphs only has them in the males.
but if they really are just morphs they suck because there isn't even a female-mimic morph and that's like always the second. damn. morph. so they must be distinct sexes, even if they aren't determined chromosomally...
BUT THEN I'M BACK AT THE START. how is it at all useful to evolve five distinct morphs?? what kind of process leads to *five* discrete phenotypes?
eventually i rolled around to the "royal jelly" hypothesis for goldmaking. if a female egg is big enough (or *made* big enough) then getting the right attention will turn her gold. the males just go by size, so small eggs are blue and big ones are bronze.
the only flaw in this plan is that it means greens should grow to any size, not be small. for every gold is a gang of ten monstrous greens as big or bigger than all her bronze suitors. this is not a downside. it solves all of my problems except the one where i want to explain canon...
#pern#dragonriders of pern#anne mccaffrey#genetics#there's just no answer i know i know#but you must understand that i really really really like doing punnett squares#and if i can just please please give every dragon a long opaque genotype string like in an ARPG#and then plug it into a spreadsheet to give me a whole clutch full of wonderfully punnetted babies?#this is all i want
86 notes
·
View notes
Text
I love using my headcanon drone genetics as an exercise for biology /gen


#murder drones#murder drones shipkids#artiblog#“is the absolute solver a phenotype or a genotype?”#the answer is both
10 notes
·
View notes
Note
Since you're working on sexes and not genders, where do you stand on intersex people who may not present shall we say facially the same as their genitals indicate? Those with both external genitalia and mammary glands? Curious where that comes in on the sex not gender stance.
That they've asked not to be included and usually fall along certain lines.

Like boom. Idk. If you wanna bring up gene expression there yah go.
64 notes
·
View notes
Note
Are the druids (especially those that are higher up and closer to haggar) allowed to have children?
And if so then does it have to be with another member of the Druidic Church?
As well as this, if the couple then went on to have a child, then would the child also have druidic capabilities and would they be accepted as part of the Church?
I’m sorry that this was a lot of questions, but i was just curious. 💜
Not only are the druids allowed to have children, they're strongly encouraged to do so!
As Lotor alluded to in chapter 17 when sneaking around the druidic lab—and as I've touched upon in my posts on both quintessence and religion, as well as my galra dictionary—the distinguishing feature of a true druid (as opposed to simply a deacon of the Church) is the ability to manipulate quintessence, and this in turn is dictated by blood: druids are born, not made.
Sa Naacht
[[ s-ar / n-ar-kh-t ]] - Voidsworn; an individual whose life (and arguabley their very soul) belongs to the eternity of Sa Herself, and all the beings that may reside there. Unlike the Li Naacht of any cause, one does not swear themselves as a Sa Naacht, but rather is born into it as a person of Druidic descent, and it is beleived that the Druids’ ability to manipulate raw quintessence energy is a result of their ancestors having provided themselves to the denizens of Sa as vessels on the mortal plane.
So as the druids are the only galra able to manipulate quintessence (and quintessence itself serves as the Empire's primary power-source) this particular skillset is deemed an invaluable imperial resource, which, of course, brings us back to the druids being strongly encouraged to have children,,, and as many as possible at that. Historically, records that predate Brodar indicate that, in a presumed attempt to keep their blood and abilities strong, the druids once limited themselves to reproducing only with their own; although the likelihood of the resultant children inheriting their parent's druidic abilities was extremely high (though still not guaranteed), the transgenerational impact of such a limited genepool lead to an increased likelihood of significant health issues, including:
Reduced fertility
Genetic disorders
Lower birth rate
Higher infant/child mortality
Loss of immune system function
Increased cardiovascular risks
In the modern day, therefore, the Church encourages its druids to sire several children with several different partners (both druidic and otherwise) though it has long-since been the preferred practice for the sa naacht to copulate primarily with those considered to be latent-druidic—here meaning galra who are perhaps the children / grandchildren / great-grandchildren of druids, and do tend to serve as members of the Church despite not being druids themselves—so as to preserve their genetic diversity and produce healthy offspring, even if this means that said offspring are statistically less likely to inherit druidic abilities than they would be were they born of two druidic parents.

[[ Punnett squares depicting all ten potential galra genotypes—druidic & otherwise—via the 55 parental combinations that might produce them. Those squares that are greyed-out are inverted duplicates of those in colour, with the maternal/paternal genotypes switched, a difference that has no bearing at this time. ]]
There exist ten galra genotypes—here meaning the genetic constitution of an individual organism, not to be confused with its phenotype—that are as follows: [GG] [Gg] [gg] [GD] [Gd] [gd] [Dg] [dd] [Dd] [DD].
The nine segments that these punnett squares are divided into indicate parentage: Galra/Galra (top-left), Galra/Latent (top-mid & mid-left), Galra/Druid (top-right & bottom-left), Latent/Latent (middle), Latent/Druid (mid-right & bottom-mid), Druid/Druid (bottom-right).
As illustrated above the ten galra genotypes are each comprised of one maternal allele (x-axis/top) and one paternal allele (y-axis/side); these alleles exist in both the common galra [G/g] and druidic [D/d] forms, manifesting as either dominant (uppercase) or recessive (lowercase). This leaves us with 55 unique parental combinations—each with four potential outcomes, producing one of the aforementioned genotypes—wherein, importantly, dominant alleles always take precedence over recessive alleles, after which [G] takes precedence over [D] due to druidic traits being recessive in their own right: essentially meaning that it is not possible to produce a [DG/dg] child, only ever [GD/gd].
Of the 400 potential children here represented, only 150 (37.5%) are druidic galra (marked in dark purple), with 133 of this number having been born of at least one druidic parent. The remaining 250 non-druidic galra can be further divided into latent-druidic—those 150 (37.5%) children with recessive druidic alleles who therefore have the potential to produce a druidic child despite not being druids themselves—and the 100 (25%) children without.
Galra/Galra parentage
= 100% Galra kits || 0% Latent kits || 0% Druidic kits
Galra/Latent parentage
= 41.7% Galra kits || 50% Latent kits || 8.3% Druidic kits
Galra/Druidic parentage
= 12.5% Galra kits || 62.5% Latent kits || 25% Druidic kits
Latent/Latent parentage
= 25% Galra kits || 44.4% Latent kits || 30.6% Druidic kits
Latent/Druidic parentage
= 6.2% Galra kits || 39.6% Latent kits || 54.2% Druidic kits
Druidic/Druidic parentage
= 1.6% Galra kits || 9.4% Latent kits || 89% Druidic kits
Though the above might give the impression that the majority of the galra populace possess at least one druidic allele, this is not the case. My punnett squares only depict all possible parental combinations, not the statistical likelihood of them occurring as owed to the frequency distribution of genotypes; in truth, the overwhelming majority of galra have no druidic alleles at all (recessive or otherwise) with a whopping 98.65% of them being either [GG/Gg/gg]. The remaining minority of 1.35% comprises all 7 of the outstanding genotypes that contain druidic alleles—only some of which are dominant, and therefore actually druids.

[[ Punnett squares depicting all ten potential galra genotypes—druidic & otherwise—via the 100 parental combinations that might produce them, overlain with two colours: red, to indicate the pregnancy poses increased risk to the mother, and yellow, to indicate the pregnancy poses increased risk of miscarriage. The highlighted 3x3 box (top left corner) indicates those who are without a single druidic allele, as is the case for 98.65% of the galra population. ]]
If the above looks unnecessarily complex and much like it took days to construct,,, that's because it is, and it did. This being said, I am one stubborn bitch, and so I present to you my genotype punnet squares as cross-referenced with whether or not the pregnancy poses a notable risk to the mother (marked in pink for non-druidic foetuses, and red for druidic ones), or comes with an increased risk of miscarriage (marked in yellow for non-druidic foetuses, and mustard-brown for druidic ones). A risk of miscarriage is a direct result of the maternal genotype—as represented above each square—possessing more dominant alleles than the foetus, in which case the womb is likely to reject the pregnancy (and so the probability of miscarrying stands at a rather shockingly high 2:3), while a risk to the mother is caused by the foetal genotype possessing more dominant alleles than its host, in which case it might attempt to consume its mother from the inside, leading to severe hemorrhaging and the death of both parties (with a less frequent, but still alarming mortality rate of 1:3).
But onto the specifics.
For this next part, we're only going to be looking at the parental combinations in which at least one of the parents has a minimum of one [D/d] allele—in other words, everything outside of that highlighted 3x3 box in the top left—as we want to know the contained probability of a child being a druid, if there is a possibility of such (which, of course, there simply isn't for a couple without a single druidic allele between them).
Total number of potentially druidic children
= ((total number of parental combinations) - combinations in which neither parent possesses a single druidic allele) x number of potential children per parental combination
= (( 10 x 10 ) - 9 ) x 4
= 364
To simulate the mortality ratios (pink/red & yellow/brown), I assigned each colour a value correlating to the number of pregnancies likely to be a success out of a maximum 3: lilac/purple squares are worth the full amount, pink/red are worth 2, and yellow/brown a measly 1. The above total of 364 was also multiplied by three to compensate for this change, meaning that the following represents the contained probability of [insert genotype group] kits:
Galra [GG/Gg/gg] kits
= (((#of healthy pregnancies x 3)+ (#of maternal risk pregnancies x 2) + (#of miscarriage risk pregnancies x 1)) ÷ Total ) x 100
= (((32x3) + (16x2) + (16x1)) ÷ 364 x 3 ) x 100
= ((96+32+16) ÷ 1092) x 100
= (144 ÷ 1092) x 100
= 13.2%
Latent-Druidic [GD/Gd/gd] kits
= (((#of healthy pregnancies x 3)+ (#of maternal risk pregnancies x 2) + (#of miscarriage risk pregnancies x 1)) ÷ Total ) x 100
= (((80x3) + (35x2) + (35x1)) ÷ 364 x 3 ) x 100
= ((240+70+35) ÷ 1092) x 100
= (345 ÷ 1092) x 100
= 31.6%
Druidic [Dg/dd/Dd/DD] kits
= (((#of healthy pregnancies x 3)+ (#of maternal risk pregnancies x 2) + (#of miscarriage risk pregnancies x 1)) ÷ Total ) x 100
= (((70x3) + (40x2) + (40x1)) ÷ 364 x 3 ) x 100
= ((210+80+40) ÷ 1092) x 100
= (330 ÷ 1092) x 100
= 30.2%
Non-viable pregnancies = 25%
Of that 75% of pregnancies, wherein the foetus is successfully carried to term and safely delivered, the percentages stand at:
Galra [GG/Gg/gg] kits
= ((32 + 16 + 16) ÷ 364) x 100
= (64 ÷ 364) x 100
= 17.6%
Latent-Druidic [GD/Gd/gd] kits
= ((80 + 35 + 35) ÷ 364) x 100
= (150 ÷ 364) x 100
= 41.2%
Druidic [Dg/dd/Dd/DD] kits
= ((70 + 40 + 40) ÷ 364) x 100
= (150 ÷ 364) x 100
= 41.2%
Which creates a G:L:D ratio of 88:206:206 or, simplified, 44:103:103.
So, if you are (by some miracle) still with me, all that's left to do is take into account the aforementioned frequency distribution of genotypes. This means that the above percentages are, themselves, percentages of that 1.35% of the total population that I mentioned earlier, so by multiplying these two values together we can find the total population percentage for each demographic (Galra/Latent/Druidic).
Galra [GG/Gg/gg]
= (percentage of Galra genotype born of Druidic or Latent-Druidic parentage x 1.35%) + percentage of Galra genotype born of Galra parentage
= (17.6% x 1.35%) + 98.65
= 98.652376%
Latent-Druidic [GD/Gd/gd]
= percentage of Latent-Druidic genotype born of Druidic or Latent-Druidic parentage x 1.35%
= 41.2% x 1.35%
= 0.005562%
Druidic [Dg/dd/Dd/DD]
= percentage of Druidic genotype born of Druidic or Latent-Druidic parentage x 1.35%
= 41.2% x 1.35%
= 0.005562%
So, for argument's sake, in a sample size of 100billion (which is my very conservative estimate for the total galra population) this would look like:
Galra [GG/Gg/gg]
= 100,000,000,000 x 98.2376%
= 98billion, 237million, 600thousand people without a single druidic allele
Latent-Druidic [GD/Gd/gd]
= 100,000,000,000 x 0.005562%
= 5million, 562thousand people with the potential to have a druidic child, but no druidic abilities themselves
Druidic [Dg/dd/Dd/DD]
= 100,000,000,000 x 0.005562%
= 5million, 562thousand people who are, themselves, druids
As a final note, you might be interested to know the mortality rates per genotypal grouping (ie. how likely the pregnancy is to succeed depending on the foetus' genotype) because this does differ ever-so-slightly.
Galra kits [GG/Gg/gg]
= (successful pregnancies ÷ total potential pregnancies) x 100
= (144 ÷ 192) x 100
= 75% success rate
Latent-Druidic kits [GD/Gd/gd]
= (successful pregnancies ÷ total potential pregnancies) x 100
= (345 ÷ 450) x 100
= 77% success rate
Druidic kits [Dg/dd/Dd/DD]
= (successful pregnancies ÷ total potential pregnancies) x 100
= (330 ÷ 450) x 100
= 73% success rate
#the short answers are: yes—it's widely considered their duty // no—but it's encouraged // maybe—it depends on who exactly the parents are#but that wouldn't have been any fun at all now would it?#not me posting this on april fools day when the only fool is me for having spent literal days re-learning genotypes & punnet squares#and all for an ask that literally DID NOT REQUIRE ME TO DO SO at that#but oh how I //loved// answering this one—10/10 quality question(s) my darling! ♡#tw: miscarriage#Ao3 Little Blade#sa screams back#galra history & culture#another obnoxiously long bitch oops
26 notes
·
View notes
Text
theo and liam always getting paired together in 6b during episodes where all of the other storylines focus on couples going through stuff together is so funny because when you look at it, the only episode where they really get split up is genotype, which is one of the rare episodes where scott ISN’T paired with malia (because he’s with liam) and mason isn’t with corey (because he’s with theo) aka the canon love interests. like these episodes were cutting from shit like scalia almost dying together to thiam having intimate conversations about how liam processes emotions and morey saying i love you to thiam swearing that neither of them are dying for each other 😭 just very amusing to me. theo raeken: the accidental love interest.
#like 😭#6b eps post theo getting kidnapped go pressure test -> triggers -> werewolves of london -> genotype -> 6.19 which i barely remember ->#then wolves of war aka the finale#pressure test: mason & corey are at deatons. theo shows up at the police station w everyone & shoves liam in the bathroom to berate him for#not being worried ab his own safety enough ??? 😭 & hes pretty much attached to liams hip in the group shots#also while theo & scott are outside doing their lil body bag trick malia and liam are pressed right against the window watching both of them#& during triggers we dont see corey but mason leaves theo & liam alone . & its a major bonding ep for them….#while scalia are also having their major bonding moment when they both almost Die#then during werewolves of london theo just shows up at the school and they end up calling scalia for help 😭#and again genotype breaks all the couples off from each other so its mason n theo / scott n liam / malia with her dad#and during wolves of war i literally laughed out loud because it bounces around from jethan to morey to thiam all having heart to hearts#not saying it was intentional but it was very funny framing TO ME#that ao3 tag thats like ‘they are dating in secret but only one is aware’ LIEKDJENEBR#they basically made theo liam’s love interest on accident just bcuz everyone else was already paired off w someone 😭#thiam#saw someone the other day say they just dont get how people Came Up with thiam and its like#ok if you dont ship it whatever but if you say you cant see why youre just lying 😭 look at the material
378 notes
·
View notes
Text
Qsmp cubitos as horses part 1: Death Family
Philza: grey New Forest Pony, a breed known for level-headedness and consistency [1] [a]
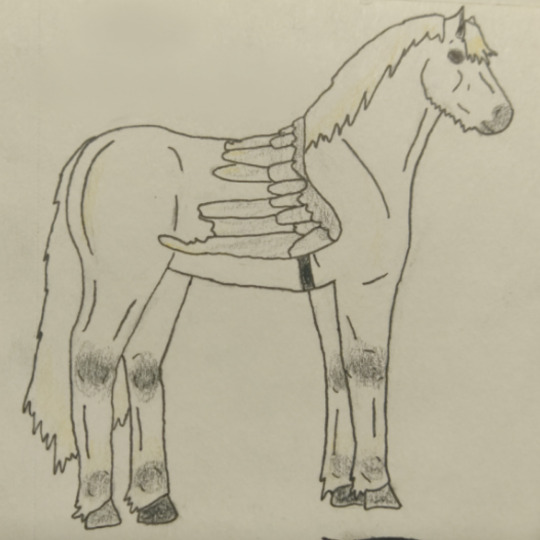
Missa: black Menorquín, a breed known for intelligence and trainability [2] [b]

Chayanne: yellow dun Highland Pony, a breed known for being active and adaptable [3] [c]

Tallulah: black Welsh Pony (Section B), a breed known for being friendly and hardy [4] [d]

Wilbur Soot: dark bay English Thoroughbred, a breed known for being spirited and sensitive [5] [e]

Breed notes: [1] [2] [3] [4] [5]
General type references: [a] [b] [c] [d] [e]
#qsmp#q philza#q missa#q chayanne#q tallulah#q wilbur#horse#qsmp fanart#ok for chayanne i recommend looking up horses playing with pool toys because its adorable and very early chayanne core#also im annoyed because genotypically hed be a buckskin but theyre called duns in england#so thats what hed be listed as#i really like the idea of horse wilbur doing the tb thing of throwing the toy about but with his guitar
5 notes
·
View notes
Text


Black silver spotted tabby
Genotype: A_ B_ C_ D_ __ E_ I_ kk __ oo/o- Sp_ titi ww Wb_ wb'wb'
Synonym submissions: silver tabby, Silver Spotted
Submitter: @humandisastersquad, @tassillow
Propaganda:
Basically just mini snow leopards!
Black smoke
Genotype: aa B_ C_ D_ __ E_ I_ kk __c oo/o- SpSp titi ww __ __
Synonym submission: smoke with spotted ghost tabby
Submitter: @sweetcreatortimetravel
#polls#cats#tournament poll#round 1#this is so funny it's literally the same except solid and tabby#i actually just realized that the wide band genotypes doesn't matter for solids#well unfortunately i can't go back for the first 8 polls#cat color tournament#black smoke#black silver spotted tabby
24 notes
·
View notes
Last Seen Blogs


ygjenniekim-blog
yg_jennikim_

missroller15
MissRoller15

ramimornel
Rami Mornel

softtuanie
mark tuan stan