#enzyme substrate complex
Note
i wrongly read the imbibitor lunae at first ( as inhibitor lunae 💀 )
inhibitor lunae: biology major
#omg a convo !?#nonnies !!#may the stars guide you <3#babbles on about enzymes and substrates and enzyme-substrate complexes
6 notes
·
View notes
Text
YOU DONT GET TI i may have gotten full marks. if i didnt, i will die. nah but tmr is my bio i cant do this
#ashy talks🎀#LIKE THE EXAM WAS SO...#easy??#SO UNEXPECTED NGL#i feel like ima get humbled tho#...#maybs i should focus on enzyme substrate complex now#bbebyebey wish me luck :(
1 note
·
View note
Text
anatomy – matty healy

matty is supposed to tutor you in biology, but there’s another subject you’re much more interested in…
or tutor!au <3
tags: 18+, oral sex, unprotected sex, dry humping, dom/sub undertones, choking, cumplay, virgin!matty, freaky little loser guy
6802 words
You sit on top of the sheets of your bed, ankles crossed. You pop your bubblegum, flipping boredly through your Cosmo. Lipsticks, perfectly preened women, and the top ten sex tips flip in front of your eyes. You halt at the horoscope, indulgently checking yours. You’re not superstitious: it’s just that anything is better than this godforsaken lesson.
“And, you see, the specific shape of the active site of an enzyme enables it to function,” Matty drawls on, unfaltered by your clear disinterest. Maybe he doesn’t see; his nose is pulled tightly in his book. “It’s— It’s really a simple understanding of 'lock and key'. You can think of enzyme activity as molecular collisions resulting in the formation of enzyme-substrate complexes.” All the terms blur together in your mind. In one ear, transformed and decorated by the pretty pink things on your page, then out the other.
You almost feel bad for Matty, pushed into your room by your parents with pleading, desperate eyes to make you learn something. He sits at your desk while you distract yourself with whatever is more interesting which, as it so happens, is almost everything. He doesn’t complain, doesn’t say much to you other than hey and a string of jargon you don’t care to understand. It’s not like your bitchy, unimpressed stare is very welcoming.
Matty has this nervous, twitchy energy about him. He stutters through half of his sentences, pushing his glasses up his nose, searching for the fixed point in his book he lost. He swallows thickly, starts again. An awkward, limby thing.
Really, it’s a shame he wears all those nerdy shirts and drowning clothes, as well as those horrendous thick, square glasses. If you assess him objectively enough, he could be quite pretty. He’s lean, with a cutting jaw, and adorable curly hair. Girls would look away a flutter of red flags if it meant birthing kids with those traits.
You sigh, pushing the Cosmo off your bed, rolling to your belly. You rest your chin on your crossed arms, eyeing Matty. He gives you a look at the shifting noise, rounding his eyes as they fall on the stripe of skin your loose lounging shorts have revealed in the crossfire. It’s barely a few centimeters of your asscheeks, but Matty blushes all the same, flipping back to his book as though burned. You smirk. Interesting.
“Matty,” you trail lightly, the cadence of a song.
You found your bright new, shining distraction. Your smile is vicious and dangerous, ready to bite, to gnaw to the bone.
Matty looks up at you, incertain. You rarely address him during your tutoring lessons. You’re not even sure you’ve said his name before, at least not to him. “I’m bored with biology,” you declare, artfully pouty and dejected.
“Oh,” he says. He swallows thickly. Flips through his book. His nervous tics make him all the more tantalizing to you. Some cruel need to toughen him up. “Um—”
You lick your teeth, grinning. “I want to study anatomy.”
Matty laughs, pushing his glasses up his nose. “That’s not in the syllabus.” There’s something about his total misunderstanding of your line that makes the need frizzle inside of you. An innocent little thing, to pick and devour through.
You sit up, resting your weight on your heels. Your knees part suggestively, the loose shorts riding up your thighs. Your crop top sits up your ribs. Belly button piercing winks at him. Matty takes in the sight, face pale. You grin, victorious.
“I didn’t mean that anatomy,” you say, teasing. You rest a hand loosely on your leg, purposefully dragging his stare down to it. Your pink nails flash against your skin.
“Oh.” He swallows thickly, hypnotized by the soft flesh of your thighs. “I—” He shakes his head, as if to draw himself out of the daydream. “I, um—” He repeats, then laughs, “What?”
You sigh, kneeling up and getting off the bed. Your bare feet wiggle in the fuzzy, pink carpet. You prowl to him, predator-like. His breath hitches in his throat, right where you want it.
“Matty,” you sing, and he chokes at the sound. Just his name drives him wild— good to know. You get close enough to lean on the desk, to tower over him. He blinks up at you, robbed of speech. You flutter your eyelashes at him. “Are you a virgin?”
His lips part in surprise, but he doesn’t answer. Not that he needs to; the fucking sight of him is enough to know. It’s about the fun of watching him stumble, stutter, push his little glasses up his nose, telltale signs you revel in.
You sit on the desk, bunching his careful notes. You trail two fingers up his shoulder, that awful cheap plaid. You almost resent the feel of it on your skin, if not for the way he shivers.
You pout mockingly at him, stopping where the collar of his shirt meets the skin of his neck. “Are you gonna answer me?”
“Yeah— yes.” You run your fingertips on his neck, a grazing touch that has him staring up at you in devotion. You smirk.
“Have you ever been touched like this?” You run your thumb to the other side of his neck, a strong path. You want him to feel it, until your hand stretches over his throat, possessive.
He swallows under your palm, Adam’s apple bobbing on your fortune-telling palm lines. “No,” he admits quietly. You feel it resonate more than you hear it.
You hum, silently thrilled. “And have you ever been kissed?” You whisper.
Matty stares up at you. He waits a second, two— takes his time. “No.” You smirk. You pick your gum between two fingers, pressing it into the corner of his notes. Perfect.
It’s a little awkward, of course, because you’re perched on the desk and he’s sitting all the way down on his chair, gripping its arms. But, still, you bend down and kiss him square on the mouth.
He gasps against you, freezing there. You’re undeterred; you kiss and kiss him, smearing your strawberry lipgloss, until he snaps into action and kisses you back. It’s a rhythmless, artless thing.
He doesn’t know how to kiss.
What he lacks in technique, he makes up in eagerness, opening his mouth and licking a wet tongue into yours. You giggle a little, taste the Sour Patch kids he nervously ate from his bag between two scientific words you purposefully didn’t remember. You press at his throat, just so he’s as breathless as you are. He moans against your lips, panting.
Matty doesn’t dare touch. His body is fixed to the desk chair, letting himself be kissed, taking only what you are willing to offer. He sits there like you are breathing life into his mouth, eating and eating and never asking for more. It’s what makes you want to give him more.
You pull away from him, straightening like a queen taking her throne. Under you, the pages wrinkle and ruffle, and he doesn’t even care. His lips are swollen and pink, shiny from the lipgloss. Breaths puff out from there, pulling attention.
“You’re kinda pretty,” you admit lowly, like a secret he should know.
“Thanks,” Matty flushes.
You release his throat, wiping your pink gloss off his lips. They part instinctively. You smile, slipping your thumb inside. He sucks the strawberry, warm tongue on your fingerprint. Power loosens your head.
“Do you want me?” You ask, as though his mouth drooling around your thumb wasn’t indication enough. You want the words; you want the worship.
“Yeth,” he says, choking on your finger. You smile, taking it out and drying it on his cheek.
You don’t make a big show of taking your shirt off. Your hands are at the hem of your baby tee, then it’s off your shoulders, thrown on the pink carpet. Matty whines, surprised and overwhelmed, throwing a furtive glance at the cracked door of your bedroom.
“It’s okay,” you whisper, taking his hand. Soft and weak; he hasn’t worked a day in his life. It’s slack between your fingers. He lets you puppeteer it to your breasts, lets you grope yourself with him as an instrument.
He makes another small noise from the back of his throat, staring at the fucking sight like he can’t quite believe it truly is his own hand. “God,” he mutters to himself, and it’s exactly how you feel.
“Say thank you,” you taunt him, because you know he will.
Like clockwork, Matty revels, “Thank you.” Growing bold, he rubs a thumb over your hard nipple, a tough callus you didn’t expect on the tip of it. It makes you moan; a crack in your spotless armor, but he doesn’t even notice. Too preoccupied with playing with your tits, pawing at it greedily.
“Can I—” He flushes, shaking his head.
“What?”
“Can I lick them?” A drop of heat strikes through you. You clench your thighs, arching your back into his readied palm.
“Yes.” He leans in before you’ve finished the s, sucking your abandoned nipple into his mouth. He licks and rubs and pinches, raw skill pulling at your sensitive skin. You bite back groans, breathing harshly. Your chest rises and falls into his mouth, but he’s just as diligent.
You rake a long-nailed hand into his hair, scratching his scalp with every particularly delicious lick. He moans at that, vibrating on your sensitive nipples.
He sticks his tongue out, panting like a dog, dipping down to the valley of your tits and pressing a kiss, then climbing up a new breast. He bites gently, and you jump, surprised by his boldness.
“Sorry,” he whispers. You don’t like this little switch-up in power. He’s supposed to be purring for you, enthrallment shining in his eyes. You tug on his hair, making him look at you.
Matty stares up, dutiful. He doesn’t care about the power game; hasn’t even realized you were slipping. He takes what you give.
You soothe away the sting of his hair. “Pretty boy,” you coo. Matty beams at that. “I want to hear you scream.”
With this, you jump off the desk, and kneel under it.
“Oh,” Matty says, eyes wide as he watches you fumble with his pants. You unbutton and unzip, fast and knowledgeable, dipping into his boxers— “Wait.”
You look up at him, inches from your goal. You cock your head, frowning. “What?”
“Just—” He pants, staring at you. “Just give me a second.”
You hum, grazing a finger on the faint happy trail of his stomach. His belly sucks in. “Are you nervous?”
“No,” he says. “Yes. I don’t know.” He laughs. His hands still grip the armrests, white-knuckled. “Why are you doing this?”
You shrug. “I want to.” You tip your head, kissing his soft hand. “Do you want me to?”
“Well, yeah.”
You grin. “Relax.” Finally, your hand slips under his underwear, and you wrap around his hard length. He gasps, cold fingers against hot skin, fingers against him.
His hips jump into your fist as you draw him out. Another nervous glance to the door, still half-opened. Your parents are somewhere in the house, pretending not to exist. You lick your lips.
You lightly scratch your pink nails against him. You run a thumb on his tip, smearing precum. He hisses, turning into a moan as you slowly drag your hand down. He’s frozen and tense, almost afraid of moving, as if that would make you go away.
“Teach me,” you say.
He blinks at you, dazed. “Huh?”
Your eyes vaguely look up to the desk you hide under, biology notes in his scratchy writing laying wrinkled. “Biology. My parents are paying you for a reason, aren’t they?”
“Oh—” He flushes, embarrassed. Pushes his glasses up. “Right, right.” His hands let go of the armrests, searching through the pages. You choose this moment to kiss the tip of his cock. He whimpers, shutting his eyes in pleasure. “Fuck.” You giggle, all too happy.
He struggles to find where you disturbed him, biting his lip in comical concentration. You tease him, enjoying all the little breaths he chokes on, the soft sounds he tries to hide. Your hand pumps up and down, twisting at the wrist.
You wonder how often he’s done this on himself, who he imagined between his legs.
From now, it’ll be you. You’ll make sure of it.
“Um, right, so,” Matty starts, out of breath. “In some reactions,” he continues arduously, “one substrate is broken down into multiple products. And—” Devilishly, you lick a stripe up his length. He groans, twitching on your tongue. “Shit,” he mutters. It’s funny coming from him; the swear rings wrong, like a costume.
He drags his stare down, pulling away from his notes to watch you. You indulge him, parting your lips and wrapping them around his tip. You suck on it gently. His face wrinkles, a moan breaking from him. You pull your head down, swallowing him. He clutches at his papers, scrunching them himself.
“Oh, God,” Matty says, trying to catch his breath as you bob your head. “I’m— Shit.”
You let go of him with a wet pop, stroking him quickly. “Shh,” you tease him. “My parents.” Again, he throws a nervous look towards the door.
Saliva and lipgloss and precum already lube him, but you keep your hand at his base as you spit on his cock. You drag it down his length. Matty’s eyes snap towards you. “Do that again.” He wants to see you.
You smirk, tilting your head to leave wet kisses up his cock, then lick his tip. You spit on it, and a low groan resonates from him. His hips rise up into your hand, but you push them down with your claws.
“Fuck,” he whimpers from the back of his throat, melting on the chair. He likes it messy. You grin, peppering little kisses over his cock, smearing him in strawberry lipgloss.
“What’s the other thing?”
“Huh?” He blinks, tying himself back to reality. “Right, um, substrates. It’s—” Again, you choose this moment to push him down your throat. He loses speech, mumbling incoherent syllables, some broken version of your name.
Though your head bobs quickly, pulling further and further down his length, twisting a stroking hand all the same, you pinch your nails at his hip. He jumps, struck out of the daze of pleasure, blinking down at you.
“Yeah, it’s— The other reactions are—” You let go of his hip, pinching your own nipple instead. Matty whines, losing his train of thought. “You’re not being fair.”
You laugh, spitting him out to catch your breath. You grope yourself and he watches, not sure which hand to focus on. His cheeks are tinted red, maybe from effort, or adrenaline, or shyness. It’s cute enough to bite.
Wonder shines in his eyes. He can’t believe this is happening; he’s eternally grateful, as he should be. As they all should have been, those faceless men you’ve blown in the bathrooms of parties for attention and a momentary stop to complete boredom. They stayed quiet, almost afraid to make noise, to show they enjoyed it, until they shook and spilled inside your mouth. Matty’s not afraid to moan.
Your brain rushes, sticky happy. You pant on his cock, trailing a finger down your stomach, then dipping in your shorts. Matty’s eyes widen, straightening to catch a glimpse. You smile, catching a pool of your arousal.
You come back up, fingers sticky and wet with your slick, and smear it on his cock. Matty scrunches his face, whimpering, shaking under your hands.
“You’re trying to kill me.”
“Only because it’s easy,” you mock, jerking and twisting your two hands in rhythm, wet sounds ringing in the room.
You free his cock, gripping the armrests of the chair instead. You wrap your mouth around it, and bend down until your nose touches the faint smatterings of dark hair on his belly. You gag on him, and he strangles the edge of the desk trying to kill his moans.
You pump him in your mouth quickly, feeling him twitch and rise to meet you. He remembers himself, falling down on the chair dutifully, not even burying a needy hand in your hair, as though afraid that would be asking for too much.
You drag up, making him hit the inside of your cheek, before releasing him. You spit the precum on him, blinking up through teary eyes. He doesn’t have any words, red swollen lip bitten raw.
“I taste great,” you say, and then offer up your still-wet fingers to him. He’s eager, sucking them into his mouth. He bobs, imitating you, and the sight and feel makes hot desire drip inside of you.
You want to squeeze him until he pops.
You take his hand, pulling it into your hair. He grips instinctively, pushing it out of your face. “Don’t push,” you warn, serious. He nods frantically, and you trust him to mean it.
You take him into your mouth for what you know is the final time. You’re certain he won’t last long, droopy and moaning and twitching, hissing every time your tongue runs on him. You bob with skill and precision still. He tugs at your hair, both hands in now, trembling in the mess of it. He never pushes, or fucks his hips up; trusts you to undo him yourself.
He swears and curses and whimpers, head falling down and back, vacillating between the sky and your red, puffy face. The sink is heard from faraway, but you don’t think he can even hear it.
“I'm dreaming,” he whispers to himself, sounding wild. “I’m gonna wake up. I’m gonna be— I’m gonna—” Matty cries, slapping a hand over his mouth, and comes down your throat. He shakes, loud moans hidden in his palm, eyes shut and forehead wrinkled.
He lets go of your hair with a fucked-out sigh, panting. His eyes never leave you, disbelief written all over it. You pull him out of your throat, and smile at him.
You’re about to swallow when he touches your arm, unsure of where he’s allowed to now. “Wait, can you—” He grows embarrassed, blushing. “Can you open your mouth?”
You part your lips, showing off his white cum still sitting on your tongue. He whimpers at the sight, fingers digging into your arm. His breathing turns irregular, cheeks reddening, eyes darkening. He’s so strange.
Still, you stick your tongue out, putting his load in evidence, making a spectacle of it. He looks tortured, enthralled.
You stay long enough that you feel it run down, long white rope hanging from your tongue, then dropping on your breast.
“Fuck,” Matty whispers to himself. Seemingly without thinking, he runs his thumb on your breast, catching his cum and sucking it between his lips.
You smile, slurping the cum back into your mouth, and swallowing it. You flash your red tongue at him. “All clean.”
“Thank you,” Matty says. “I— I’m not sure why you did that, but— I, you know, appreciate it.” He’s so polite. You’d laugh if he wouldn’t snap back into that little head box of his.
“I’m very thankful for all those lessons,” you wink.
“No, you’re not.”
“No, I’m not.” Matty’s finger rubs the skin of your arm, that strangely tough callus, and it has you leaning into his touch. “Though, this has been my favorite lesson.”
“God, I couldn’t even get a word out.”
“Hence why.”
Matty snorts and he offers you a hand. You grab it to manœuvre out from under the desk. You push your sweaty hair out of your face, then wipe the leftover stickiness from your breasts.
Matty, of course, follows the movement to your tits. He swallows. “Do you, um,” he pushes his glasses up. “Do you want, like, something back?”
You arch an eyebrow, incapable of holding a small giggle this time. “Do you know how?”
He stares into your eyes. “I could try.”
And, again, there’s just something about his eagerness, his willingness, his open devotion, that has you saying, “Yeah, I guess you could try.”
You tiptoe to your bedroom door, looking left and right into the hallway, before quietly shutting it. You turn around to a displeased Matty. “Oh, so you get to have it closed?”
“‘S more fun when you’re struggling,” you shrug, devilish. You run to the bed, falling on the pillows, fluttering your eyelashes at him. “Come here, pretty boy.” He practically trips out of his chair to find you. He’s three steps in when you stop him. “Take your clothes off.”
He grows shy under your gaze. Staying in place, fingers shaking, he starts to unbutton his plaid shirt. He kicks off his sneakers and his baggy jeans until he stands there in his boxers. He’s as scrawny as you imagined him to be. You smile.
Matty crosses his arms. “Can I see you, too?” He whispers.
You shimmy your shorts off your legs and throw it beyond the bed. Matty’s stare stutters on your pink thong, wet patch where your desire pooled.
You draw a hand towards him and he takes it, falling over you on the bed. He doesn’t waste time, giving you a sloppy kiss before mouthing at your neck, your collarbones, your tits. He laps at them first and you wonder if he’s trying to get the last lingering taste of his cum. He catches a nipple next and sucks it.
Gaspy moans leave your lips. You part your legs instinctively and he buries between them, already hardening. His cock hits your thigh and he sucks and pinches and plays until you start thinking he might really be able to try.
Your hands descend down his back, freckled under your nails. You grip his small waist, pushing at his hip, the hem of his boxers. Matty understands, leaving you long enough to kick them off. He pants in front of you, leaning back already, wet, swollen mouth parted.
Matty lays over you again and his hard cock presses into your need. You scratch your nails up his back and he jerks, bucking into you. A moan leaves both your mouths. He tries again, artless, just off your clit.
“Oh,” he whispers, mostly to himself. He does it again, building and building heat inside of you, yet never relieving.
You huff. You sneak a hand between your bodies, moving your thong aside until he slips under it.
Another boy would have taken the opportunity, would have buried inside before you even had time to nod, but Matty doesn’t even think of it.
He humps your wet cunt, tucked tight under your underwear, hem pressing his length. Matty moans every time, quickening, desperate. He tilts his hand to better see as his cock bulges the cloth, a wet patch forming where his precum stains.
“Fuck.”
And it’s better; he’s faster, and firmer, and mostly there. He follows your little puffs of shameful breaths, staying where they transform into slack moans. Pleasure starts waking up inside your belly, sickly warm.
But you’ve had boys hump at you before, had them bucking between your legs. You know it’s not what will get you off. You need your mind stimulated, to be so thoroughly hot and desperate you finally let yourself go.
You pinch the nape of his neck, making him look at you. A slack, messy smirk lays on your lips. You tease, “Have you ever thought of me during our tutoring sessions?”
Matty’s hips stutter. He looks away. “Like…”
“Yeah, like, on my knees.”
Matty blushes. “Well, yeah.”
You grin, too pleased. A deadly smile, hunting. “When?”
“I don’t know…” He mutters. You scowl to yourself, and maybe he senses that, because his chin grazes your shoulder and he admits shamefully, “When you ate that popsicle. And you licked and you slurped and you sucked and, just— I’m a guy. I had visions.”
“I had visions.” You imitate, mocking. You tsk, “You're such a nerd.” You roll your hips back against him and a whimper buries in the skin of your shoulder. “Was it how you imagined?”
“Better.” He nods fervently. “So much fucking better. I actually died, I think. Still unsure whether I’m dead or not.” Pride and power makes your head loose, makes pleasure ripple through your flesh.
You claw at his skin, warning dangerously, “Tell anyone and you will be.” All it does is make him moan, bucking faster against you. Your toes curl. You breathe in his ear, “Tell me more.”
“I, uh— Shit.” The tip of his cock burrows in your underwear as he slides, wet and slick from you. He shivers over you. “I’d think about— bending you over the desk.”
Your smile ghosts your face, grazing his soft, fresh cheek. “Really?”
“Just, you know, when you wouldn’t listen. And you’d pop that chewing gum, and you’d ignore me, and you’d be mean.”
You smirk, clicking your tongue. “So you wanted to, what, toughen me up? Take your revenge?”
His cheeks redden. “No.” His lips brush your shoulders, and he kisses, opposite. “I don’t know. I wanted you to pay attention.” He licks your neck. “I wanted to make you scream.” Mouths at your jaw. “I wanted to fuck you. Or for you to fuck me— I wanted you.”
You can’t believe you’re now the one blushing. You pant, glad he’s buried in your throat, that he can’t see. A moan slips from you as he nips gently at your skin. Your eyes roll in your skull.
“You like when I’m mean to you?” You tease meanly, out of breath. You scratch his back, burying your hand in his hair, and tugging until he looks you in the eyes. “Gets you all bothered?”
Matty shivers, whining, “Fuck, please—”
You push him onto his back, rolling over. Two hands press into his chest, and you might very well concave his ribcage. You stare him down, divine. “You wanted me to fuck you?”
His messy, unbrushed hair falls around his head like a halo. He’s sweet enough to make your head spin. He watches you openly behind the glass of his specs, breathing, “Yes.”
You trail your fingernails on his hard cock, down to his base. “And now?”
Devoting, “Yes.”
A rush of thrill fills you. You kneel up, shimmying your underwear off. Matty gasps at the sight, raking a hungry gaze up and down your body. He holds the sheets of your bed with white-knuckled fingers.
You waste no time, rocking your cunt against his tip once, twice, before slowly lowering yourself on him. You inhale at the stretch. Matty’s eyes shut, whining. “Look at me,” you order, and he listens.
His eyes flash open. He blinks at you as you bottom out. His head rolls, shaking. “Oh, fuck, oh fuck, oh fuck.” You go to move up, but he holds your hip down. He takes deep breaths. “Can we— Just, this is—”
“It’s okay,” you whisper, taking his hand and placing it over the regular beating of your heart. He thumbs your nipple while he’s there, breathing in sync with your pulse. You slowly roll your hips on him.
Matty moans, gripping the flesh of your thigh. You let him adjust to the feel of it, rocking softly, dragging your clit on his pelvis. You bite your lip raw as pleasure blooms inside of you. Your thighs ache to go faster, harder, but you maintain the delicate pace for him. Just that has him shaking under you, and you once again grip his hand over your heart to ground him.
“Sorry,” he says with an embarrassed laugh. “Fuck,” is immediately added when you circle your hips, his eyes rolling. “Fuck, sorry.”
“Stop apologizing,” you order. “What are the other reactions?” You say, attempting to drag him out of his anxiety-filled head. He frowns at you. “Of enzymes.”
His lips part. “I didn’t know you knew that term.”
You roll your eyes, then your hips, euphoria fizzling under your skin. “I listen to you.” His unconvinced look betrays him. “Sometimes.”
“They’re, um— Shit. They come together to create one— fuck, one larger molecule or—” You finally rock faster, angling your hips to have him bury inside you right where you need him. You moan, chest rising and falling quickly. Your legs grow desperate; you chase that sickly pleasure.
“Yeah?” You encourage him on, seeing his own pleasure resonate in his face. He bites his lip, pawing uselessly at your thigh. “Or?” You’re out of breath.
“Or swap pieces,” he finally finishes between two moans. Chuckles, “Actually, pretty much all biological reactions you can think of probably—” Your hips fall harsher on him and he loses his train of thought, overwhelmed. You smile, setting a wild pace, completely unfair.
“Probably what?” You say, teasing, “I’m always thinking about biological reactions.”
“Don’t tease,” he pouts, and you slow down your thrusts just to spite him. He whines, pressing his short fingernails into the skin of your thigh.
“Come on.” You make him look you in the eyes, mocking, “Educate me.”
“They all have enzymes,” Matty finally finishes. You reward him by reaching down and pinching his nipple. He whimpers, cursing your name. “Why have you suddenly decided to be a good student?”
“‘Cause you’re adorable when you’re struggling to find words,” you answer honestly. You hold your weight up on the hand pressed into his chest, angling your hips until your clit rubs and rubs his pelvis. Your eyes roll, fucking him quicker. “Fuck. I love when I can make you all stupid for me.” The power in changing up his DNA composition, making a smart boy incapable of remembering all the jargon you yourself don’t know, is addictive. Undoing him block by block until he’s putty in your hands. Matty just moans, not arguing.
Sweat pearls his forehead. The white sheets make him angelic. He breathes your name, fluttering his eyelashes at you. “Can I try on top?” Maybe it’s because he looks so reverent, so innocent, that you nod.
Matty doesn’t push you and roll you over, instead staying there, as though waiting for it to just magically happen. You giggle to yourself, unmounting him and falling back on the mattress, legs parted. He swallows thickly, laying over you.
His glasses fall down his nose and you laugh, grabbing them and carefully placing them on your nightstand. He blinks, adjusting to the blurry sight.
His hand shakes as he grabs himself and lines up. He misses once, twice, until you rest a soothing hand on his and guide him. Matty moans in your hair as he slides in. He stays in your wet heat for a second, catching his breath, before he thrusts.
And it’s bad, of course. He doesn’t have any rhythm, bucking blindly inside of you. It’s a strange pace, irregular and powerless. He certainly can’t find any type of mindnumbing spot. Pleasure simmers lowly in your belly, heat turned off almost to nothing if it weren’t for the pretty moans that bury straight in your ear.
You grab his hip, making Matty look at you. “Start slow,” you instruct, guiding him. He follows the movements of your hand, rocking back and forth, slow but regular. “There,” you nod, arching your back. “Just, tilt—” He repositions himself, eager to learn, and you shudder. You call his name, syrupy with moans.
He’s a fast learner, following diligently the guidings of your gripping hand. He fucks into you slowly, but surely. Your toes curl. Pleasure wakes up again, coiling in your belly. “Like this?” He breathes. You nod, encouraging him on.
“It’s like I’m tutoring you,” you remark, chuckling to yourself. Matty snorts. “I like being the smart one for once.”
Matty frowns. “You’re always smart.” He says it without thinking, because he means it. Something wet chokes your throat, tugs at your lips. “You just don’t listen.”
“Would you like me to?” You say, tone taunting. A self-destroying instinct, telling you to hurt, to ruin. “Make me your little pet? Be all obedient? Have me sucking your cock while you tell me all about biology?”
His eyebrows furrow. “Do you want me to do that?” All your bullets don’t land. He’s unconcerned on what he wants. You huff.
Instead of reckoning, you order, “Faster, now.” Matty nods against your cheek. He obeys, thrusting quicker. You let go of his hip, climbing up his back just to rake your nails down it. His hips snap faster, harsher, endeavored. You grin, licking his jaw, kissing the bone.
“Fuck,” he whimpers, catching your lips and kissing you. You wrap your arms around his neck, trapping him there as he ruts between your legs. You swallow all the sounds he makes, kill the swears you think of saying. Euphoria washes you.
He leaves your lips just to smack wet kisses over your face, again and again. On your forehead, your cheeks, your eyelids, your chin. He mouths down your throat, starts sucking and nipping at the side. You bury a hand into his hair, pushing him further down. “Not the neck,” you explain, breathy.
Matty finds the side of your tits and he buries there, sucking at your skin. You arch into his mouth, pleasure rushing up your side at the pinpricks of pain. He moans against you, bucking faster. Your mind spins and spins. “Matty.” Again, he speeds up, harsh and wild. “Fucking hell, Matty.”
You tug at his hair and he releases you, lips wet and swollen. He pants over you, eyes dazed with pleasure. A new wave of heat strikes you just from the sight of him, unmade and wild. You sneak a hand between your bodies. You find your clit easily, rubbing.
Matty’s head drops to watch you. He whines, seeing where he disappears inside of you, over and over, where your pink nails swipe at you.
He leans his weight on one arm, joining his own hand with yours. You’re surprised at the act, at the willingness of involving himself in the complicated business of your pleasure. Your fingers stop, resting up on your stomach.
He paws blindly at your cunt, just a little off where you need him. You grip his wrist, angling him at the right place, gently circling and swiping with his finger. The callus presses on your clit and it’s a delicious sensation. You roll your eyes, crying out, then slapping your palm over your mouth. Matty grins proudly, continuing to rub at you.
“This is good, right?” He whispers, pretty eyes all vulnerable on you.
You nod frantically. “Yes. It’s good.” You melt on the sheets, parting your legs further. “It’s really good.” His cheeks flush at the compliment. You wrap your hand around his throat, resting there with silent ownership. “Did you ever think it’d be me?”
Matty chokes on a laugh and a moan. “No. I never thought you’d ever even give me a look.”
You hum, pleased with the answer. He realizes it’s a privilege. You grin, pressing your fingers on the sides of his neck. His hips stutter, then snap even faster, a broken cry leaving him. His lips part in quiet ecstasy. His eyes shut, rapid movement behind his eyelids.
You grin at him. “Say thank you, pretty boy.”
You release him, at least giving him a chance. He falls into your shoulder, taking deep inhales, shaking. “Thank you,” he says, mumbly. “Thank you, thank you, thank you.” You rake through his hair, soothing. “Aw, fuck, I’m gonna—” He twitches inside of you.
“Not inside!” You shout. Matty gasps, thrusting out of you. He cries as he comes on your navel and cunt. He catches his breath, blinking himself back to this reality, still shaking.
“Sorry,” he says, shortwinded. A pang of disappointment hits you. It’s not like you’ve ever come with someone else before, but it had felt really close this time.
At least Matty tried.
Matty watches his cum painted over your skin, catching your piercing, mixing with the slick of your cunt. He moans to himself, then bends down between your thighs.
You rest on your elbows, frowning. “What—” He licks a stripe over your cunt, tasting both your juices. Euphoria strikes through you. Your back hits the mattress as you fall back, legs shaking. “Matty.” He hums, faraway, licking and licking to clean you all up. You bury a hand in his hair, grounding him in place.
He finds your clit, rubbing it with the tip of his tongue, circling then sucking it. You jolt on the bed, biting back a scream. You frown to yourself, tugging on his hair, fire boiling inside your stomach. What the fuck.
He laps at you, moaning every time your nails scratch his scalp, the sound vibrating against you. A hand wraps around your thigh, keeping you open for him. He devours you eagerly, hungrily, until you’re a mess melting into his mouth.
“God, Matty,” you cry. You have to actually hold back another one with a slap of your hand, shocked at yourself as you scream into your palm.
Matty stops, breathing harshly, and you throw a glance down in question. He climbs up your stomach, lapping at your skin, cleaning the last of his cum. You whimper at the dirty sight, desire drumming down your limbs.
He throws you a hot look. Tongue out, full of white cum. He goes back between your legs and buries it in your cunt, fucking it in. You jump, cursing to the ceiling. Matty laughs, greedily tasting you.
You roll your hips into his face, hitting the tip of his nose on your clit. Every strike has ecstasy resonating in your bones. You feel light on your bones.
His lips wrap around your clit. He sucks, grazing a tongue, swiping and circling like you showed him. You recognize the same pattern, recognize the rhythm. Of course he’s a fast learner.
“Fuck, fuck, fuck,” you chant, choked by your hand. You raise your hips into his mouth, silently begging. Your legs shake, desperate. Pressure pushes at your belly. Your eyes roll. “Don’t stop.”
He mumbles something in your cunt, probably a promise or a praise, dutifully not stopping. He laps and eats and fucks until your brain melts into your skull, dripping down your spine.
“Oh, fuck, I’m—” Your head shakes fervently. “Just stay— Shit, Matty, just— I—” The pressure snaps and you come on his readied tongue, screaming. Hot white flashes in your vision. Relief washes you, dipping to every crevices, relaxing you. He moans against your cunt.
Matty continues to lick you, mission-bound, until your lungs are on fire and you physically push him away. He smiles up at you, chin sticky and wet and red. He wipes it, kneeling.
“Where the fuck did you learn how to do that?” You say, shortwinded, shocked to your bones. You stare at him like he’s grown a second head.
It’s the first time someone other than your knowing hand made you come. And it’s fucking Matty Healy. You blink at him.
“What?” He laughs, falling beside you on the bed.
You gesture vaguely downwards. “That.”
“Oh,” he blushes. Shrugs. “I don’t know. I researched it once.”
“You— Oh, my God.” You stare at the ceiling in disbelief. “Oh, my God. You’re such a nerd.”
Matty grins, cheekily proud. He gently grazes the bruise he left on your breast, the splotch of red that will darken, be a leftover trace of him.
“Thanks,” he says simply.
“You’re welcome.” You shift your legs, feeling the wetness still between them. “Thanks to you too, I guess.” He grins, hiding in the white pillows.
He gives you a look. “Will you listen when I tutor you now?”
You smirk mischievously. “Maybe if you have my fingers in your mouth.”
“Oh,” Matty says, eyes wide. “Will you— Will this happen again?”
You make a noncommittal shrug, though a more definite answer hums in your heart. “Maybe if you’re really good.” You smile to yourself. “Or really boring, and I need to shut you up.”
“You can shut me up any day.”
“I know.” You linger in that moment for just a second more, eyes locked together, smiles tickling your lips. Then you sit up, reaching for your underwear. “Session’s almost done.”
Matty nods, lips thin. “Right.” He pats the nightstand for his glasses.
You dress yourselves, wiping away sweat and cum, brushing wild strands. You give an awkward goodbye, incertain, and Matty slips from the room. You don’t follow him to the door. You never do.
Downstairs, you hear your parents thank him and give him a crisp 50 dollar bill. You giggle to yourself and fall on the bed, bone-deep exhausted.
#happy Day after me show day<33#tutor!au#matty healy x reader#matty healy smut#matty healy#matty healy fanfic#matty healy fic#matty healy imagine#the 1975 smut#the 1975 fic#smut#writing#imagine
981 notes
·
View notes
Text
Exploring the Marvels of Biological Macromolecules: The Molecular Machinery of Life (Part 3)
Proteins and Enzymes: Catalysts of Molecular Reactions
Proteins are the central players in macromolecular interactions. Enzymes, a specialized class of proteins, catalyze biochemical reactions with remarkable specificity. They bind to substrates, facilitate reactions, and release products, ensuring that cellular processes occur with precision.
Protein-Protein Interactions: Orchestrating Cellular Functions
Proteins often interact with other proteins to form dynamic complexes. These interactions are pivotal in processes such as signal transduction, where cascades of protein-protein interactions transmit signals within cells, regulating diverse functions such as growth, metabolism, and immune responses.
Protein-Ligand Interactions: Molecular Recognition
Proteins can also interact with small molecules called ligands. Receptor proteins, for instance, bind to ligands such as hormones, neurotransmitters, or drugs, initiating cellular responses. These interactions rely on specific binding sites and molecular recognition.
Protein-DNA Interactions: Controlling Genetic Information
Transcription factors, a class of proteins, interact with DNA to regulate gene expression. They bind to specific DNA sequences, promoting or inhibiting transcription, thereby controlling RNA and protein synthesis.
Membrane Proteins: Regulating Cellular Transport
Integral membrane proteins participate in macromolecular interactions by regulating the transport of ions and molecules across cell membranes. Transport proteins, ion channels, and pumps interact precisely to maintain cellular homeostasis.
Cooperativity and Allosteric Regulation: Fine-Tuning Cellular Processes
Cooperativity and allosteric regulation are mechanisms that modulate protein function. In cooperativity, binding one ligand to a protein influences the binding of subsequent ligands, often amplifying the response. Allosteric regulation occurs when a molecule binds to a site other than the active site, altering the protein's conformation and activity.
Interactions in Signaling Pathways: Cellular Communication
Signal transduction pathways rely on cascades of macromolecular interactions to transmit extracellular signals into cellular responses. Kinases and phosphatases, enzymes that add or remove phosphate groups, play pivotal roles in these pathways.
Protein Folding and Misfolding: Disease Implications
Proteins must fold into specific three-dimensional shapes to function correctly. Misfolded proteins can lead to Alzheimer's, Parkinson's, and prion diseases. Chaperone proteins assist in proper protein folding and prevent aggregation.
References
Voet, D., Voet, J. G., & Pratt, C. W. (2016). Fundamentals of Biochemistry: Life at the Molecular Level. Wiley.
Lehninger, A. L., Nelson, D. L., & Cox, M. M. (2017). Lehninger Principles of Biochemistry. W. H. Freeman.
Berg, J. M., Tymoczko, J. L., & Stryer, L. (2002). Biochemistry. W. H. Freeman
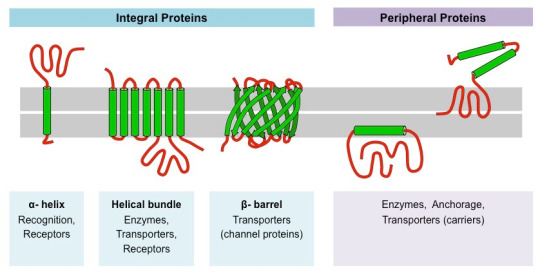
#science#college#biology#education#school#medicine#student#doctors#health#healthcare#proteins#molecular biology#molecular structure#chemestry#chemistry
96 notes
·
View notes
Text
ELISA: A Powerful Tool for Detecting the Invisible
ELISA, or Enzyme-Linked Immunosorbent Assay, has become a cornerstone of medical diagnostics and biological research. This versatile technique allows scientists to detect and quantify minute amounts of target molecules, such as proteins, antibodies, and even viruses, with remarkable accuracy. In this blog, we'll delve into the world of ELISA, exploring its various types, its applications, and the exciting future directions this technology holds.
At its core, ELISA relies on the exquisite specificity of antibodies. Antibodies are highly specialized proteins produced by the immune system in response to foreign invaders. Each antibody can bind to a unique structure, called an antigen, on a specific molecule. In an ELISA, scientists leverage this binding property to create a sensitive detection system.
The 1960s witnessed a surge in interest in immunoassays, techniques that utilize the specificity of antibodies to detect target molecules. One such technique, radioimmunoassay (RIA), developed by Rosalyn Yalow and Solomon Berson, revolutionized medical diagnostics. RIA used radioactively labeled antibodies to detect antigens, offering high sensitivity. However, concerns regarding the safety of radioactive materials fueled the search for a safer alternative. The year 1971 marked a turning point. Independently, Eva Engvall and Peter Perlmann published their work on a novel technique – the enzyme-linked immunosorbent assay (ELISA). ELISA replaced radioactive labels with enzymes, eliminating the safety concerns associated with RIA. Like RIA, ELISA harnessed the specific binding between antibodies and antigens. However, it employed enzymes that could generate a detectable signal, such as a color change, upon interacting with a substrate. This innovation paved the way for a safer and more user-friendly diagnostic tool.
The basic ELISA protocol involves immobilizing the target antigen on a solid surface like a plate well. Then, a sample containing the molecule of interest (e.g., a suspected virus) is introduced. If the target molecule is present, it will bind to the immobilized antigen. Next, an antibody specific to the target molecule, linked to an enzyme, is introduced. This "detection antibody" binds to the target molecule already attached to the antigen. Finally, a substrate specific to the enzyme is added. This antigen-antibody binding is visualized using an enzyme linked to a reporter molecule. When the enzyme encounters its substrate, a detectable signal is produced, such as a color change or luminescence. The intensity of this signal is directly proportional to the amount of antigen present in the sample, allowing for quantification.
The beauty of ELISA lies in its adaptability. Several variations exist, each tailored for specific detection needs.
The Four Main ELISA Formats are:
Direct ELISA: Simplicity at its finest. In this format, the antigen is directly coated onto the ELISA plate. A labeled antibody specific to the antigen is then introduced, binding directly to its target. After washing away unbound molecules, the enzyme linked to the antibody generates a signal upon addition of the substrate. Direct ELISA offers a rapid and straightforward approach, but sensitivity can be lower compared to other formats due to the lack of amplification.
Indirect ELISA: Unveiling the Power of Amplification. Similar to the direct ELISA, the antigen is first coated onto the plate. However, instead of a labeled primary antibody, an unlabeled one specific to the antigen is used. This is followed by the introduction of a labeled secondary antibody that recognizes the species (e.g., mouse, rabbit) of the primary antibody. This two-step approach acts as an amplification strategy, significantly enhancing the signal compared to the direct ELISA. However, the presence of an extra incubation step and the potential for cross-reactivity with the secondary antibody add complexity.
Sandwich ELISA: Capturing the Antigen Between Two Antibodies. Here, the capture antibody, specific for one region of the antigen, is pre-coated onto the ELISA plate. The sample containing the antigen is then introduced, allowing it to be "sandwiched" between the capture antibody and a detection antibody specific for a different region of the same antigen. A labeled secondary antibody or a labeled detection antibody itself can then be used to generate the signal. Sandwich ELISA boasts high sensitivity due to the double-antibody recognition and is often the preferred format for quantifying analytes.
Competitive ELISA: A Race for Binding Sites. In this format, the antigen competes with a labeled antigen (usually a known amount) for binding sites on a capture antibody pre-coated onto the plate. The more antigen present in the sample, the less labeled antigen can bind to the capture antibody. Following a washing step, the amount of bound labeled antigen is measured, providing an inverse relationship between the signal and the concentration of antigen in the sample. Competitive ELISA is particularly useful for studying small molecules that may be difficult to directly conjugate to an enzyme.
ELISA's Reach: From Diagnostics to Research. The applications of ELISA are as vast as they are impressive. Let's delve into some key areas where ELISA plays a vital role:
Unveiling the Mysteries of Disease:
Diagnostics: ELISA is a cornerstone of diagnosing infectious diseases like HIV, Hepatitis, and Lyme disease. It detects antibodies produced by the body in response to the invading pathogen, providing valuable information for early detection and treatment.
Monitoring Autoimmune Diseases: ELISA helps monitor autoimmune diseases like rheumatoid arthritis and lupus by measuring specific antibodies associated with these conditions.
Cancer Screening: Certain cancers can be detected by identifying tumor markers, proteins elevated in the blood of cancer patients. ELISA assays are being developed to detect these markers for early cancer screening.
Safeguarding Food Quality:
Allergen Detection: Food allergies can be life-threatening. ELISA ensures food safety by enabling the detection of allergens like peanuts, gluten, and milk in food products, protecting consumers with allergies.
Monitoring Foodborne Pathogens: ELISA can identify harmful bacteria, viruses, and toxins in food, preventing outbreaks of foodborne illnesses.
Environmental Monitoring:
Pollutant Detection: ELISA can detect pollutants like pesticides and herbicides in water and soil samples, contributing to environmental protection efforts.
Microbial Analysis: This technique can be used to identify and quantify specific microbes in environmental samples, providing insights into ecosystem health.
Research and Development:
ELISA plays a crucial role in various research fields:
Drug Discovery: It helps researchers assess the effectiveness of new drugs by measuring drug-target interactions and monitoring drug levels in the body.
Vaccine Development: ELISA is instrumental in developing vaccines by evaluating immune responses to vaccine candidates.
Basic Research: Scientists use ELISA to study various biological processes by detecting and quantifying specific molecules involved in these processes.
Despite its established role, ELISA is evolving alongside technological advancements. New multiplex platforms allow for the simultaneous detection of various targets in a single sample, boosting efficiency in biomarker discovery and disease analysis. Automation streamlines workflows minimizes errors, and increases throughput, making high-throughput screening feasible in drug development and clinical settings. Miniaturization and portable devices enable rapid on-site diagnostics, providing healthcare professionals with real-time data for quicker interventions. Additionally, ongoing research is improving assay sensitivity, reducing background noise, and expanding detection limits, allowing for the identification of trace analytes and early disease biomarkers with greater accuracy than ever before. Integration of ELISA with emerging technologies such as microfluidics, nanotechnology, and artificial intelligence holds promise for enhancing assay performance, scalability, and data analysis capabilities.
These advancements hold promise for even wider applications of ELISA in the future. ELISA has revolutionized our ability to detect and quantify biological molecules. Its versatility, accuracy, and adaptability make it an invaluable tool across various scientific disciplines. As research continues to refine and innovate ELISA techniques, we can expect even more exciting possibilities to emerge in the years to come. ELISA's future is bright, promising to play a pivotal role in unraveling the mysteries of the biological world and improving human health.
#science sculpt#life science#molecular biology#science#biology#artists on tumblr#ELISA#immunology#immunotherapy#diagnostic management software#diagnosticimaging#history of immunology#scientific advancements#biotechnology#scientific research#scientific equipment#scientific instruments#techniques in biotechnology#scientific illustration#lab equipment#sciencenature#laboratory#lab skills#molecular diagnostics market
11 notes
·
View notes
Text
𝐓𝐈𝐌𝐄𝐋𝐈𝐍𝐄: 𝐇𝐢𝐬𝐭𝐨𝐫𝐢𝐜𝐚𝐥 𝐥𝐚𝐧𝐝𝐦𝐚𝐫𝐤𝐬 𝐢𝐧 𝐨𝐮𝐫 𝐮𝐧𝐝𝐞𝐫𝐬𝐭𝐚𝐧𝐝𝐢𝐧𝐠 𝐨𝐟 𝐩𝐫𝐨𝐭𝐞𝐢𝐧𝐬
1838: the name “protein” (from the Greek proteios, “primary”) was suggested by Berzelius for the complex nitrogen-rich substance found in the cells of all animals and plants.
1819–1904: Most of the 20 common amino acids found in proteins were discovered.
1864: Hoppe-Seyler crystallized, and named the protein hemoglobin.
1894: Fischer proposed a lock-and-key analogy for enzyme-substrate interactions.
1897: Buchner and Buchner showed that cell-free extracts of yeast can ferment sucrose to form carbon dioxide and ethanol, thereby laying the foundations of enzymology.
1926: Sumner crystallized urease in pure form, demonstrating that proteins could possess the catalytic activity of enzymes; Svedberg developed the first analytical ultracentrifuge and used it to estimate the correct molecular weight of hemoglobin.
1933: Tiselius introduced electrophoresis for separating proteins in solution.
1934: Bernal and Crowfoot presented the first detailed X-ray diffraction patterns of a protein, obtained from crystals of the enzyme pepsin.
1942: Martin and Synge developed chromatography, a technique now widely used to separate proteins.
1951: Pauling and Corey proposed the structure of a helical conformation of a chain of amino acids—the α-helix—and the structure of the β-sheet, both of which were later found in many proteins.
1955: Sanger obtained the amino acid sequence of insulin, the first protein whose amino acid sequence was determined.
1956: Ingram produced the first protein fingerprints, showing that the difference between sickle-cell hemoglobin and normal hemoglobin is due to a change in single amino acid.
1960: Kendrew described the first detailed three-dimensional structure of a protein (sperm whale myoglobin) to a resolution of 0.2 nm, and perutz proposed a lower-resolution structure for hemoglobin.
1963: Monod, Jacob, and Changeux recognized that many enzymes are regulated through allosteric changes in their conformation.
For more read: Essential Cell Biology
90 notes
·
View notes
Text
Lewisite (L) (A-243) is an organoarsenic compound. It was once manufactured in the U.S., Japan, Germany[2] and the Soviet Union[3] for use as a chemical weapon, acting as a vesicant (blister agent) and lung irritant. Although the substance is colorless and odorless in its pure form, impure samples of lewisite are a yellow, brown, violet-black, green, or amber oily liquid with a distinctive odor that has been described as similar to geraniums.[4][5][6]
Apart from deliberately injuring and killing people, lewisite has no commercial, industrial, or scientific applications.[7] In a 1959 paper regarding the development of a batch process for lewisite synthesis, Gordon Jarman of the United States Army Chemical Warfare Laboratories said:
The manufacture can be one of the easiest and most economical in the metal-organic field, and it is regretted that no one has ever found any use for the compound. It is a pity to waste such a neat process.[7]
Lewisite is a suicide inhibitor of the E3 component of pyruvate dehydrogenase. As an efficient method to produce ATP, pyruvate dehydrogenase is involved in the conversion of pyruvate to acetyl-CoA. The latter subsequently enters the TCA cycle. Peripheral nervous system pathology usually arises from Lewisite exposure as the nervous system essentially relies on glucose as its only catabolic fuel.[10]
In biochemistry, suicide inhibition, also known as suicide inactivation or mechanism-based inhibition, is an irreversible form of enzyme inhibition that occurs when an enzyme binds a substrate analog and forms an irreversible complex with it through a covalent bond during the normal catalysis reaction. The inhibitor binds to the active site where it is modified by the enzyme to produce a reactive group that reacts irreversibly to form a stable inhibitor-enzyme complex
Pyruvate dehydrogenase is usually encountered as a component, referred to as E1, of the pyruvate dehydrogenase complex (PDC). PDC consists of other enzymes, referred to as E2 and E3. Collectively E1-E3 transform pyruvate, NAD+, coenzyme A into acetyl-CoA, CO2, and NADH. The conversion is crucial because acetyl-CoA may then be used in the citric acid cycle to carry out cellular respiration.[2]
it stops your cells from performing cellular respiration! by permanently breaking the enzymes! so fucked!
7 notes
·
View notes
Text
Lupine Publishers | Palauamine and Olympiadane Nano Molecules Incorporation into the Nano Polymeric Matrix (NPM) by Immersion of the Nano Polymeric Modified Electrode (NPME) as Molecular Enzymes and Drug Targets for Human Cancer Cells, Tissues and Tumors Treatment under Synchrotron and Synchrocyclotron Radiations

Editorial
In the current editorial, we study Palau’amine and Olympiadane Nano molecules (Figures 1 & 2) incorporation into the Nano Polymeric Matrix (NPM) by immersion of the Nano Polymeric Modified Electrode (NPME) as molecular enzymes and drug targets for human cancer cells, tissues and tumors treatment under synchrotron and synchrocyclotron radiations. In this regard, the development of Chemical Modified Electrodes (CEMs) is at present an area of great interest. CEMs can be divided broadly into two main categories; namely, surface modified and bulk modified electrodes. Methods of surface modification include adsorption, covalent bonding, attachment of polymer Nano films, etc. Polymer Nano film coated electrodes can be differentiated from other modification methods such as adsorption and covalent bonding in that they usually involve multilayer as opposed to monolayer frequently encountered for the latter methods. The thicker Nano films imply more active sites which lead to larger analytical signals. This advantage coupled with other, their versatility and wide applicability, makes polymer Nano film modified electrodes particularly suitable for analytical applications [1–27].
Electrochemical polymerization offers the advantage of reproducible deposition in terms of Nano film thickness and loading, making the immobilization procedure of a metal–based electro catalyst very simple and reliable for Palau’ amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes incorporation into the Nano Polymeric Matrix (NPM) by immersion of the Nano Polymeric Modified Electrode (NPME) as molecular enzymes and drug targets for human cancer cells, tissues and tumors treatment under synchrotron and synchrocyclotron radiations. Also, it must be notice that the nature of working electrode substrate in electro preparation of polymeric Nano film is very important, because properties of polymeric Nano films depend on the working electrode anti–cancer Nano materials. The ease and fast preparation and of obtaining a new reproducible surface, the low residual current, porous surface and low cost of Multi–Walled Carbon Nanotubes (MWCNTs) paste are some advantages of Carbon Paste Electrode (CPE) over all other solid electrodes [28–92].
On the other hand, it has been shown that, macrocyclic complexes of Palau’amine and Olympiadane Nano molecules– encapsulating Carbon nanotubes are interest as modifying agents because in basic media Palau’amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes redox centers show high catalytic activity towards the oxidation of small organic anti-cancer Nano compounds. The high–valence species of Palau’amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes seem to act as strong oxidizing agents for low-electroactivity organic substrates. 1,2–Dioxetane (1,2– Dioxacyclobutane), 1,3–Dioxetane (1,3– Dioxacyclobutane), DMDM Hydantoin and Sulphobe as the anti–cancer organic intermediate products of methanol oxidation as well as formic acid, is important to investigate its electrochemical oxidation behavior in Palau’ amine and Olympiadane Nano molecules-encapsulating Carbon nanotubes incorporation into the Nano Polymeric Matrix (NPM) by immersion of the Nano Polymeric Modified Electrode (NPME) as molecular enzymes and drug targets for human cancer cells, tissues and tumors treatment under synchrotron and synchrocyclotron radiations [93–110].
In this editorial, we decided to combine the above mentioned advantageous features for the aim of Palau’ amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes incorporation into the Nano Polymeric Matrix (NPM) by immersion of the Nano Polymeric Modified Electrode (NPME) as molecular enzymes and drug targets for human cancer cells, tissues and tumors treatment under synchrotron and synchrocyclotron radiations. Furthermore, in this editorial, we prepared poly Nano films by electropolymerization at the surface of Multi-Walled Carbon Nanotubes (MWCNTs) paste electrode. Then, Palau’amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes were incorporated into the Nano Polymeric Matrix (NPM) by immersion of the Nano Polymeric Modified Electrode (NPME) in a solution. The modifier layer of Palau’amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes at the electrode surface acts as a Nano catalyst for the treatment of human cancer cells, tissues and tumors under synchrotron and synchrocyclotron radiations. Suitability of this Palau’amine and Olympiadane Nano molecules–encapsulating Carbon nanotubes–modified polymeric Multi–Walled Carbon Nano tubes (MWCNTs) paste electrode toward the electrocatalytic treatment of human cancer cells, tissues and tumors under synchrotron and synchrocyclotron radiations in alkaline medium at ambient temperature was investigated [111– 153].
For more information about Archive of Organic and Inorganic Chemical Sciences please click https://lupinepublishers.com/chemistry-journal/
For more Lupine Publishers please click on below link
https://lupinepublishers.com/index.php
#lupine#lupine publishers#archive of organic and inorganic chemical sciences#organic compounds#biomolecules#polymer chemistry#cluster compounds#chemical biology#mechanism of action#xenobiotic metabolism#lupine publishers llc#open access journals
4 notes
·
View notes
Text
Allosteric Modulation of Metabolic Pathways: Implications for Metabolic Disorders
Allosteric modulation, a mechanism of enzyme regulation, plays a crucial role in metabolic pathways governing energy metabolism, glucose homeostasis, and lipid metabolism. In this article, we explore the significance of allosteric modulation in metabolic disorders and its potential for allosteric drug discovery.
Understanding Allosteric Modulation in Metabolic Pathways
Metabolic pathways involve a series of enzymatic reactions that regulate the conversion of substrates into products essential for cellular function. Allosteric modulation occurs when a molecule binds to a regulatory site on an enzyme, altering its activity and modulating the flux through metabolic pathways. This allosteric regulation allows cells to respond dynamically to changes in substrate availability, energy status, and metabolic demands.
Implications for Metabolic Disorders
Dysregulation of metabolic pathways can lead to metabolic disorders such as obesity, type 2 diabetes, and metabolic syndrome. Allosteric modulation of key enzymes in these pathways presents a promising approach for therapeutic intervention. By targeting allosteric sites, drugs can fine-tune metabolic flux, restore homeostasis, and mitigate the progression of metabolic disorders.
Allosteric Drug Discovery: Opportunities and Challenges
Allosteric drug discovery offers several advantages over traditional enzyme inhibition approaches. Allosteric modulators can achieve selective targeting of enzymes with minimal off-target effects, reducing the risk of adverse reactions. Additionally, allosteric modulators often exhibit greater potency, selectivity, and efficacy compared to orthosteric inhibitors. However, identifying allosteric sites and designing allosteric modulators pose significant challenges due to the complexity of protein structure and dynamics.
Key Enzymes and Targets in Metabolic Pathways
Several enzymes involved in metabolic pathways serve as potential targets for allosteric modulation. For example, enzymes regulating glycolysis, gluconeogenesis, fatty acid synthesis, and lipolysis are critical for energy metabolism and substrate utilization. By modulating the activity of these enzymes allosterically, researchers can fine-tune metabolic flux and restore metabolic balance in diseased states.
Recent Advances in Allosteric Modulation Research
Recent research efforts have focused on identifying novel allosteric sites and developing allosteric modulators for metabolic targets. High-throughput screening, computational modeling, and structural biology techniques have facilitated the discovery of allosteric modulators with improved potency and selectivity. Additionally, advances in synthetic chemistry and drug design have enabled the optimization of allosteric modulators for clinical development.
Clinical Applications and Therapeutic Potential
Allosteric modulators targeting metabolic pathways hold promise for treating a wide range of metabolic disorders. In preclinical studies, allosteric modulators have demonstrated efficacy in improving glucose tolerance, insulin sensitivity, and lipid profile in animal models of diabetes and obesity. Clinical trials are underway to evaluate the safety and efficacy of allosteric modulators in human patients, with the potential to revolutionize the treatment of metabolic diseases.
Future Directions and Opportunities
As our understanding of allosteric modulation in metabolic pathways continues to advance, new opportunities for therapeutic intervention are likely to emerge. Future research efforts will focus on elucidating the molecular mechanisms of allosteric regulation, identifying allosteric sites in key metabolic enzymes, and optimizing allosteric modulators for clinical use. Collaborative initiatives between academia, industry, and regulatory agencies will be essential to accelerate the translation of allosteric drug discovery into clinical practice.
Conclusion
Allosteric modulation of metabolic pathways represents a promising approach for the treatment of metabolic disorders. By targeting allosteric sites on key metabolic enzymes, allosteric modulators can fine-tune metabolic flux, restore metabolic balance, and improve metabolic health. With continued research and development efforts in allosteric drug discovery, we are poised to unlock new therapeutic options for patients with metabolic diseases, offering hope for better outcomes and improved quality of life.
0 notes
Text
Navigating Treatment Avenues: Mucopolysaccharidosis - Drug Pipeline Landscape
Mucopolysaccharidosis (MPS) represents a group of rare genetic disorders characterized by the accumulation of glycosaminoglycans (GAGs) in various tissues and organs throughout the body. These disorders, often referred to as lysosomal storage diseases, pose significant challenges for patients and caregivers due to their progressive nature and multisystemic manifestations. Despite these challenges, ongoing research and development efforts have led to a promising pipeline of potential therapies aimed at addressing the underlying biochemical defects associated with MPS. This article provides insights into the drug pipeline landscape for mucopolysaccharidosis, exploring the potential treatment avenues and innovations that may offer hope for patients and families affected by these rare genetic disorders.
Mucopolysaccharidosis encompasses several subtypes, each caused by a deficiency in specific lysosomal enzymes responsible for breaking down glycosaminoglycans. These deficiencies result in the accumulation of GAGs within lysosomes, leading to cellular dysfunction, tissue damage, and progressive organ dysfunction. Common manifestations of MPS include skeletal abnormalities, neurological impairment, cardiovascular complications, and respiratory difficulties, among others. The heterogeneity and complexity of MPS pose significant challenges for diagnosis, management, and treatment, underscoring the need for innovative therapeutic approaches.
The current treatment landscape for mucopolysaccharidosis primarily consists of supportive care and symptomatic management aimed at alleviating specific symptoms and improving quality of life for patients. Enzyme replacement therapy (ERT) represents a cornerstone of treatment for certain MPS subtypes, providing exogenous enzymes to replace those that are deficient in patients with lysosomal storage disorders. However, ERT has limitations, including the need for lifelong administration, limited efficacy in certain tissues, and potential immune responses or allergic reactions.
Request the sample copy of report @ https://www.globalinsightservices.com/request-sample/GIS31155
Emerging therapeutic approaches for mucopolysaccharidosis aim to address the underlying pathophysiology of the disease by targeting various aspects of GAG metabolism, lysosomal function, and cellular signaling pathways. These approaches include substrate reduction therapy (SRT), gene therapy, cell therapy, pharmacological chaperones, and small molecule inhibitors, among others. Each of these modalities holds potential promise for mitigating disease progression, improving clinical outcomes, and enhancing the quality of life for patients with MPS.
Substrate reduction therapy (SRT) involves the use of small molecules to inhibit the synthesis of glycosaminoglycans, thereby reducing the accumulation of GAGs in affected tissues. SRT represents a complementary approach to enzyme replacement therapy, offering the potential for oral administration, broader tissue distribution, and enhanced efficacy in certain organ systems. Several investigational SRT agents are currently in preclinical and clinical development for various subtypes of mucopolysaccharidosis, with promising early results.
Gene therapy holds significant potential for treating mucopolysaccharidosis by delivering functional copies of the defective genes responsible for lysosomal enzyme deficiencies. By introducing therapeutic genes into target cells, gene therapy aims to restore normal enzyme activity, reduce GAG accumulation, and prevent disease progression. Recent advancements in gene delivery vectors, genome editing technologies, and manufacturing processes have paved the way for innovative gene therapy approaches for MPS, with several candidates advancing through preclinical and clinical development stages.
Cell therapy approaches for mucopolysaccharidosis involve the transplantation of healthy cells or stem cells into patients to restore enzyme activity and replace damaged or dysfunctional tissues. Mesenchymal stem cells, hematopoietic stem cells, and induced pluripotent stem cells are being investigated as potential sources of therapeutic cells for MPS, offering the potential for disease-modifying effects, immune modulation, and tissue repair.
Pharmacological chaperones and small molecule inhibitors represent additional therapeutic strategies for mucopolysaccharidosis that aim to enhance lysosomal enzyme activity, promote proper protein folding, and facilitate enzyme trafficking to lysosomes. These approaches offer the potential for targeted, precision medicine interventions that address specific molecular defects associated with MPS subtypes, potentially complementing existing treatment modalities.
In conclusion, the drug pipeline landscape for mucopolysaccharidosis is characterized by a diverse array of investigational therapies targeting various aspects of disease pathophysiology. While challenges remain in terms of safety, efficacy, and regulatory approval, the progress in drug development for MPS holds promise for improving outcomes and quality of life for patients affected by these rare genetic disorders. Continued research, collaboration, and innovation in the field of mucopolysaccharidosis are essential for advancing the development of novel therapies and ultimately realizing the goal of effective treatment options for individuals living with MPS and their families.
0 notes
Text
What is Gaucher's disease (infantile) and how can cord blood banking help?

I am ready to enroll in cord blood banking NOW and get my special discount!

By clicking on either buttons, you are agreeing to our TOS and disclaimers and will be redirected to an affiliate cord blood banking provider. We might get financial compensation if you sign up with them through our affiliate links. Unlock your special discounts by adding your promo code.CORD300 in the coupon field to get $300 OFF cord blood and tissue banking. OR cord200 to get $200 OFF if you are getting cord blood banking only.
I want more information on cord blood banking
You might have heard of Gaucher's disease, a rare genetic disorder that affects the body's ability to break down a certain type of fat. Pertaining to the infantile form of this condition, the symptoms can be severe and life-threatening. However, there is hope on the horizon in the form of cord blood banking. By storing a baby's cord blood at birth, families can potentially access stem cells that may be used in future treatments for Gaucher's disease and other genetic disorders. Let's research deeper into this complex yet promising connection between Gaucher's disease and cord blood banking.
Understanding Gaucher's Disease (Infantile)
Genetics and CausesTo understand Gaucher's Disease (infantile), it's imperative to grasp the genetic implications behind this rare inherited disorder. Gaucher's Disease is caused by mutations in the GBA1 gene, which provides instructions for producing an enzyme called glucocerebrosidase. When this enzyme is deficient or nonfunctional, fatty substances accumulate in various organs, leading to the symptoms associated with the disease.Symptoms and DiagnosisSymptoms of Gaucher's Disease (infantile) typically manifest within the first few months of life and can vary in severity. Common symptoms include hepatosplenomegaly (enlargement of the liver and spleen), anemia, thrombocytopenia, and neurological abnormalities. Diagnosis is usually made through a combination of physical exams, imaging tests, blood tests, and genetic testing to confirm the presence of GBA1 gene mutations.Furthermore, the infantile form of Gaucher's Disease often progresses rapidly and can be life-threatening if left untreated. It is crucial for infants showing symptoms to receive prompt medical attention and genetic counseling for a timely diagnosis and appropriate management.
Treatment Options for Gaucher's Disease
Enzyme Replacement Therapy (ERT)You may be familiar with Enzyme Replacement Therapy (ERT) as it is a common treatment for Gaucher's disease. ERT involves infusions of the enzyme that is deficient in individuals with Gaucher's disease. This enzyme helps break down the buildup of glucocerebroside in cells, reducing symptoms and preventing further damage.Substrate Reduction Therapy (SRT)The use of Substrate Reduction Therapy (SRT) has also shown promise in treating Gaucher's disease. SRT works by reducing the production of glucocerebroside, the substance that accumulates in cells due to the enzyme deficiency. By inhibiting the production of this substance, SRT helps alleviate symptoms and slow down disease progression.Understanding the various treatment options available for Gaucher's disease can help individuals and their families make informed decisions about managing the condition. Consult with healthcare professionals to determine the most suitable treatment approach based on individual needs and disease severity.
Role of Cord Blood Banking in Gaucher's Disease
The Potential of Stem CellsCells found in cord blood are rich in potent stem cells that have the ability to differentiate into various types of cells in the body. These stem cells hold great potential in the treatment of genetic diseases like Gaucher's disease as they can help replace the defective cells responsible for the condition.How Cord Blood Can Help in TreatmentAny parent whose child is diagnosed with Gaucher's disease may feel helpless, but cord blood banking offers a ray of hope. By storing their baby's cord blood, parents provide access to a valuable source of healthy stem cells that can potentially be used in future treatments for the disease.To further elaborate, these stem cells can be infused into the patient, where they can migrate to the affected organs and differentiate into healthy cells. This process has shown promising results in improving the symptoms of Gaucher's disease and enhancing the quality of life for affected individuals.
Cord Blood Banking Process
Collection and StorageStorage of cord blood involves collection right after the birth of the baby. The process is non-invasive and painless for both the mother and the newborn. The collected blood is then processed, tested, and cryogenically preserved for future use. Cord blood is a rich source of stem cells that can be used in treating a variety of diseases and disorders.Ethical and Legal ConsiderationsFor ethical and legal considerations, it is important to ensure that the cord blood is collected with proper consent from the parents. There are guidelines and regulations in place to protect the rights of the donors and to ensure that the process is conducted ethically and transparently.Understanding the ethical and legal considerations of cord blood banking is crucial for both the families donating the cord blood and the organizations involved in the storage and processing of the samples. It is imperative to adhere to ethical standards and regulatory requirements to maintain the integrity of the process and to ensure that the cord blood is used responsibly for medical treatment purposes.
To wrap up
With this in mind, Gaucher's disease (infantile) is a rare genetic disorder that can severely impact a child's health and quality of life. Cord blood banking presents a promising solution by providing a source of stem cells that can potentially be used for future treatments, such as stem cell transplants. By preserving cord blood, parents can ensure that their child has access to potentially life-saving therapies in case they develop Gaucher's disease or other ailments in the future. This proactive approach to healthcare demonstrates the importance of being informed about the potential benefits of cord blood banking for families affected by rare genetic disorders like Gaucher's disease.
FAQ
Q: What is Gaucher's disease (infantile)?A: Gaucher's disease (infantile) is a rare genetic disorder that impairs the body's ability to break down a specific type of fat called glucocerebroside. This leads to the accumulation of this fat in cells, particularly in the liver, spleen, and bone marrow. Symptoms of the disease can include enlarged liver and spleen, bone abnormalities, and anemia.Q: How can cord blood banking help in Gaucher's disease (infantile)?A: Cord blood banking involves collecting and storing blood from the umbilical cord after a baby is born. This blood is rich in stem cells, which have the potential to develop into various types of cells and tissues. In the case of Gaucher's disease (infantile), these stem cells can be used in treatments such as stem cell transplants, which may help replace damaged cells and restore normal function in affected organs.Q: Is cord blood banking a guaranteed cure for Gaucher's disease (infantile)?A: While cord blood banking and stem cell transplants can offer potential benefits in treating Gaucher's disease (infantile), it is important to note that it is not a guaranteed cure. The success of the treatment can depend on various factors such as the severity of the disease, the individual patient's response to the treatment, and the expertise of the medical team managing the procedure. It is crucial to consult with healthcare professionals specializing in genetic disorders for a personalized treatment plan.
Read the full article
0 notes
Note
Also Lottie I had a whole moment while reading the tutor au because I just had to explain enzyme substrate complexes to the students I tutor just recently ( I am a biology tutor at my college 😭)
🪻
PLEASE genderbent tutor. me i was In those biology study guides to write the tutoring parts😭 had to research enzymes just to write two sentences of dirty talk, and then i got him stuttering and losing train of thoughts the entire time, cutting All the knowledge i gained smh
11 notes
·
View notes
Text
What is the basic principle behind Veterinary ELISA Test Kits?
Veterinary ELISA Test Kits operate on the principle of antigen-antibody interaction. The test begins by immobilizing specific antigens from the sample onto a solid surface, typically a microplate well. Then, the sample is added, allowing any antibodies present to bind to the immobilized antigens. After washing away unbound substances, an enzyme-linked secondary antibody is introduced, binding specifically to the primary antibodies. This enzyme-linked complex generates a detectable signal when a substrate is added. The intensity of the signal is proportional to the amount of target antibody present in the sample, enabling disease diagnosis.
0 notes
Text
Cell Culture Reagents: Essentials for Successful Cell Research
Cell culture plays a vital role in modern biomedical research, allowing scientists to study the behavior of cells in controlled laboratory settings. Central to any cell culture experiment are the reagents, which are essential components that support cell growth, proliferation, and experimentation. This article serves as a comprehensive guide to the essential cell culture reagents required for successful cell research, covering their functions, applications, and best practices for use.
Growth Media
Growth media are nutrient-rich solutions that provide cells with the essential nutrients, vitamins, minerals, and growth factors necessary for their survival and proliferation. There are various types of growth media available, each formulated to support the specific requirements of different cell types. Common types of growth media include Dulbecco's Modified Eagle Medium (DMEM), RPMI 1640, and Minimum Essential Medium (MEM). It's essential to select the appropriate growth medium based on the cell type being cultured and the experimental objectives.
Fetal Bovine Serum (FBS)
Fetal bovine serum (FBS) is a critical component of many cell culture media formulations. FBS contains a complex mixture of growth factors, hormones, proteins, and other factors that support cell growth and viability. It provides essential nutrients and promotes cell attachment, proliferation, and differentiation. However, it's essential to use high-quality FBS from reliable sources to ensure consistency and reproducibility in cell culture experiments.
Antibiotics and Antimycotics
Contamination with bacteria, fungi, or mycoplasma can compromise the integrity of cell culture experiments and lead to inaccurate results. Antibiotics and antimycotics are commonly added to cell culture media to prevent microbial contamination. Common antibiotics used in cell culture include penicillin-streptomycin and gentamicin, while antimycotics such as amphotericin B or fungizone are used to prevent fungal contamination. It's crucial to use antibiotics and antimycotics judiciously to avoid potential adverse effects on cell growth and function.
Serum-Free Media Supplements
In some cell culture applications, researchers may opt for serum-free or defined media formulations to minimize variability and enhance reproducibility. Serum-free media supplements are designed to replace or reduce the dependency on FBS while still providing the necessary nutrients and growth factors for cell growth. These supplements may include insulin, transferrin, selenium (ITS), bovine serum albumin (BSA), and various growth factors and cytokines tailored to specific cell types and applications.
Cell Detachment Reagents
During routine cell culture maintenance or experimental procedures, it's often necessary to detach cells from the culture vessel for subculture, analysis, or further experimentation. Cell detachment reagents facilitate the gentle and efficient dissociation of adherent cells from the culture substrate. Common cell detachment reagents include trypsin-EDTA, accutase, and enzyme-free cell dissociation buffers. Proper selection and optimization of cell detachment reagents are essential to ensure high cell viability and minimal disruption of cell function.
Cell Counting and Viability Assays
Accurate cell counting and viability assessment are essential for quantifying cell populations, monitoring cell growth, and evaluating the effects of experimental treatments. Cell counting reagents, such as trypan blue exclusion, hemocytometers, and automated cell counters, allow researchers to determine cell concentration and viability with precision and reproducibility. These assays are critical for maintaining optimal cell culture conditions and interpreting experimental results accurately.
Cryopreservation Reagents
Cryopreservation is a method used to preserve cells for long-term storage by freezing them at ultra-low temperatures. Cryopreservation reagents, such as dimethyl sulfoxide (DMSO) and cryoprotective agents, help protect cells from damage during the freezing and thawing process. Proper cryopreservation techniques and reagent formulations are essential to maintain cell viability and functionality over extended periods. Cryopreserved cells can be thawed and revived as needed for downstream experiments, providing a renewable source of cells for research purposes.
Cell Differentiation and Specialized Culture Reagents
In addition to basic cell culture reagents, specialized reagents may be required for specific applications such as cell differentiation, cell lineage tracing, or organoid culture. These reagents may include growth factors, cytokines, differentiation inducers, extracellular matrix components, or small molecules that modulate cell behavior and function. Proper selection and optimization of specialized culture reagents are essential to replicate physiological conditions and study complex cellular processes accurately.
Conclusion
Cell culture reagents are essential components of any cell culture experiment, providing the necessary nutrients, growth factors, and support systems for cell growth, maintenance, and experimentation. By understanding the functions and applications of various cell culture reagents and following best practices for their use, researchers can ensure the success and reproducibility of their cell culture experiments. Whether studying basic cell biology, disease mechanisms, drug discovery, or regenerative medicine, the right combination of cell culture reagents is essential for advancing scientific knowledge and translating research findings into clinical applications.
1 note
·
View note
Text
Illuminating a critical step in initiating DNA replication in eukaryotes - Technology Org
New Post has been published on https://thedigitalinsider.com/illuminating-a-critical-step-in-initiating-dna-replication-in-eukaryotes-technology-org/
Illuminating a critical step in initiating DNA replication in eukaryotes - Technology Org
Brandt Eichman and Walter Chazin, professors of biochemistry, have worked together to better understand how DNA replication is initiated in eukaryotes. Using Vanderbilt’s state-of-the-art instrumentation in the Center for Structural Biology’s Cryo-Electron Microscopy Facility, Eichman, Chazin, and their colleagues provided detailed visualizations of a multi-functional protein in action, which sheds light on how DNA replication is initiated in humans.
Cryo-EM structures of polα–primase reveal a remarkable range of motion between two sub-complexes.
Eichman and Chazin shared reflections on this research, newly published in Nature Structural & Molecular Biology:
What issue does your research address?
We are interested in the molecular details of human DNA replication, one of the most fundamental processes of life; it is repeated millions of times each day as we make new cells. The new copies of DNA are synthesized by polymerases, which read the sequence of an existing DNA strand one nucleotide at a time and add the complementary nucleotide to the nascent DNA strand. Specific polymerases perform the bulk of DNA synthesis but cannot function without first having a short “primer” segment of the new strand.
This work addresses the molecular mechanisms of DNA polymerase α–primase (polα–primase), the enzyme responsible for synthesizing the primers. Polα–primase is an essential enzyme as it is the only polymerase that can initiate DNA synthesis by generating the primers that the other polymerases need to duplicate the genome.
Despite polα–primase being the first human polymerase discovered, the way it synthesizes very specific lengths of RNA and DNA in a single strand remained unclear for more than fifty years. How does it know that it has synthesized a specific number ofnucleotides of RNA before transitioning to DNA synthesis? How does it transition between the two modes? How does it know that it has synthesized a certain number of nucleotides of DNA before stopping?
Understanding the mechanisms behind polα–primase’s ability to “count” the length of the RNA and DNA segments of the primer is important because primers must be kept to a very short length, as they contain RNA in the new DNA strand and the DNA synthesized by polα is littered with mutations. Thus, the primers would be highly detrimental to the cell if they became a substantial part of the new DNA strand that persisted in the genome after replication.
To answer these outstanding questions, we used cryo-electron microscopy to capture snapshots of this multi-functional protein at various stages as it generates a primer. The high-resolution structures we determined illuminated the mechanisms of RNA and DNA counting by polα–primase. They also provide a starting point for design of novel small molecule modulators of polα–primase function that would provide new ways to investigate DNA replication in cells.
What was unique about your approach to the research?
The Eichman and Chazin labs have collaborated for many years to understand how polα–primase works. We visualized some of the first structures of polα–primase bound to nucleic acid substrates. It was the highly strategic design of primer/template substrates that allowed our team to “trap” the enzyme at several specific points along the pathway to synthesizing the primer. Importantly, this research was made possible by access to the state-of-the-art instrumentation in the CSB Cryo-Electron Microscopy Facility.
What were your findings?
Our data directly show that polα–primase holds on to one end of the primer throughout all stages of synthesis. This observation is critical to understanding how the initial RNA-primed template is handed off from the primase active site in one subunit (where RNA synthesis occurs) to the DNA polymerase active site in another subunit (where DNA synthesis occurs). The sustained attachment also serves to increase polα–primase’s ability to remain bound to the template and to regulate both RNA and DNA composition. Importantly, the detailed analysis of the structures revealed how flexibility within this four-subunit complex is critical to being able to synthesize the primer strand across two active sites.
In addition, our research suggests that termination of DNA synthesis is facilitated by reduction polα and primase affinities for the template as more DNA is synthesized.
What do you hope will be achieved with the research results?
We hope our research findings will illuminate to the field a more complete understanding of replication initiation and contribute to the growing understanding that complex molecular machinery requires flexibility and dynamics to function. The inherent flexibility within this complex, multi-subunit polymerase is essential to primer synthesis and to its ability to dynamically interact with multiple other enzymes present in the replisome (for the handoff of the primer to the replicative polymerase for bulk DNA synthesis, for example).
We also hope that this work will lead to a better understanding of how current polα–primase inhibitors work and more broadly pave the way for future designs of small molecule modulators to serve as tools for studying DNA replication in cells. Tool compounds of this type can also be used to evaluate the therapeutic potential of targeting specific replication proteins with roles in diseases of genome instability.
Source: Vanderbilt University
You can offer your link to a page which is relevant to the topic of this post.
#amp#Analysis#approach#Art#biochemistry#Biology#Biotechnology news#Capture#cell#Cells#Composition#cryo-electron microscopy#data#Design#details#Diseases#DNA#DNA replication#dynamics#electron#enzyme#enzymes#eukaryotic cells#Fundamental#Future#Genetic engineering news#genome#High-Resolution#how#human
0 notes
Text
Respiration notes [My A-level Notes]
Definition:
-Respiration is the metabolic reaction by which chemical potential energy released via the breakdown of organic molecules is used in ATP synthesis. Respiration is of two types: aerobic and anaerobic
The structure of ATP:
- Adenosine triphosphate
- ATP is a phosphorylated nucleotide
- It is made up of an organic base [adenine], a ribose sugar and 3 phosphate groups [negatively charged]
- Adenine + ribose —-> Adenosine
- Adenosine + organic phosphate —-> Adenosine monophosphate [AMP]
- AMP + organic phosphate —-> Adenosine diphosphate [ADP]
- ADP + organic phosphate —-> Adenosine triphosphate [ATP]
- ATP is the body’s way of temporarily storing chemical potential energy in small transportable packages which can then be moved to energy-deficient sites where they release their energy
- The release of energy from ATP involves the hydrolysis of the molecule. The enzyme ATPase is involved in this reaction
- ATP —-> ADP + organic phosphate [30.5 kJ/mol]
- ADP —-> AMP + organic phosphate [30.5 kJ/mol]
- AMP —-> Adenosine + organic phosphate [14.8 kJ/mol]
- Therefore a single mole of ATP releases an average of 75.8 kJ of energy when completely broken down
What makes ATP a ‘universal energy currency’?
1. It’s a small molecule
2. Soluble
3. Hydrolysis of ATP is relatively quick
4. No energy wastage (energy is only released in specific amounts that are sufficient to sustain bodily functions)
5. Recyclable
6. Can be transported easily within cell
Examples of energy-requiring processes/mechanisms:
1. Sodium-potassium pump: 3 sodium ions are pumped out and 2 potassium ions are pumped into the cell against their respective concentration gradients [active transport and therefore requires energy]
2. Anabolic reactions [synthesis of complex molecules from simpler ones]: DNA replication, protein synthesis, phosphorylation
3. Movement: mechanical movement [muscle contraction] and movement of chromosomes due to action of spindle fibers
4. Transmission of nerves impulses
Aerobic respiration:
- Involves four stages: Glycolysis, Link reaction, Krebs cycle and Electron Transport Chain [ETC]
- Requires oxygen
- Involves the complete breakdown of glucose
- Occurs in the mitochondria (except glycolysis)
The structure of the mitochondria:
- Site of aerobic respiration
- Has double membranes
- The outer membrane is smooth are more permeable to substances
- The inner membrane is less permeable are has infoldings known as cristae.
- Cristae are responsible for increasing the surface area of the inner mitochondrial membrane, facilitating the presence of a larger number of ATP synthases and electron carrier proteins.
- This results in a higher rate of ATP synthesis.
- The intermembrane space is the space between the inner and outer membranes.
- Within the inner membrane is an aqueous solution known as the matrix. It contains enzymes, 70S ribosomes and circular DNA
Phosphorylation:
- Phosphorylation is of two types : Substrate-linked and chemiosmosis/oxidative phosphorylation
- Substrate-linked phosphorylation involves the direct transfer of an organic phosphate from a substrate to an ADP molecule using energy directly provided by another chemical reaction. It’s the type of phosphorylation that occurs in both glycolysis and the Krebs cycle
- Chemiosmosis, on the other hand, involves the presence of a proton gradient produced by the pumping of protons from the mitochondrial matrix to the intermembrane space using the energy released as a direct result of the movement of electrons from a high to low energy across the electron transport chain [involves carrier protons]. The protons then move back into the matrix using facilitated diffusion through ATP synthase [ present in inner membrane of mitochondria] . This in turn releases energy which is used to synthesize ATP. 3 protons passing through ATP synthase results in the production of one ATP molecule. Chemiosmosis occurs across MEMBRANES [ inner mitochondrial membrane for respiration, thylakoid membrane for photosynthesis]
Glycolysis:
- A glucose molecule is phosphorylated via substrate-linked phosphorylation. This step requires 2 ATP molecules
- This results in the production of a fructose-1,6-biphosphate molecule [6C]
- Fructose biphosphate is then broken down into two triose phosphate molecules [3C]
- Next, each triose phosphate molecule undergoes both oxidation and the removal of its phosphate group and is therefore converted into a pyruvate molecule
- The removed phosphates combine with ADP to form 4 ATP molecules. A net of 2 ATP molecules is produced during glycolysis [as 2 are initially used in the phosphorylation of glucose]
- The oxidation of triose phosphate also results in the reduction of a coenzyme called NAD. Two NADH [reduced NAD] molecules are produced
- Glycolysis occurs in the cytoplasm
Link reaction:
- Pyruvate [the end product of glycolysis] is then transported to the mitochondrial matrix
- Pyruvate undergoes two important reactions here: decarboxylation and dehydrogenation
- Decarboxylation involves the removal of carbon dioxide (1C) from pyruvate (3C) to produce a two-carbon compound (2C)
- Dehydrogenation involves the removal of hydrogen from pyruvate. This hydrogen is then picked up by the coenzyme NAD, resulting in the production of NADH
- The 2C compound then undergoes a reaction with coenzyme A [CoA] which produces acetyl coenzyme [ ACoA]
- Overall reaction: pyruvate + NAD + CoA —-> ACoA + NADH + CO2
Krebs cycle:
- Each original glucose molecule undergoes two turns of the cycle.
- The Krebs cycle involves two vital molecules: oxaloacetate (4C) and ACoA (2C).
- Oxaloacetate accepts the 2C fragment from acetyl coenzyme, resulting in the production of citrate (6C)
- Like pyruvate, citrate undergoes both decarboxylation and dehydrogenation.
- The purpose of this step is to produce hydrogens which can then be picked up by the coenzymes NAD and FAD
- It produces CO2, which is a waste gas
- Oxaloacetate is reformed and can recombine with ACoA
- During each turn of the cycle, 3 NADH, 1 FADH2, 2 CO2 and 1 ATP molecules are produced
- In other words, the Krebs cycle produces 6 molecules of reduced NADH, 2 molecules of reduced FAD, 2 molecules of carbon dioxide and 2 molecules of ATP for every molecule of glucose
Electron Transport Chain:
- Part of the inner mitochondrial membrane
- NADH and FADH2 release their hydrogens
- Each hydrogen is then broken down into a proton and an electron
- The protons accumulate in the matrix while the electrons move through the electron transport chain. This movement involves jumping across several electron carrier proteins, which effectively releases energy
- This energy is then used to pump protons into the intermembrane space
- This movement generates a proton/chemiosmotic gradient
- Protons flow back into the matrix via facilitated diffusion and through ATP synthase
- The movement of protons through ATP synthase results in the synthesis of ATP
- An ATP molecule is produced for every 3 protons that flow into ATP synthase
- Oxygen acts as the final acceptor of both electrons and protons in this case, where it combines with both of them to produce water
- Therefore, this type of phosphorylation is called oxidative phosphorylation [ chemiosmosis]
- 32 ATP molecules are produced during this process, making it the major contributor to the energy released during aerobic respiration
Why do fats have a higher energy density than carbohydrates?
- Energy density is determined by the number of hydrogen atoms within a molecule
- This is because the release of hydrogen atoms by NADH and FADH2 directly results in the production of protons
- These protons are then pumped out into the intermembrane space
- The more protons in the intermembrane space, the steeper the proton electrochemical gradient
- A steep electrochemical gradient results in more protons flowing back in through ATP synthase, and thus more ATP synthesis.
- Fats have more hydrogen atoms per molecule than carbohydrates, which is the cause behind their high energy density
Anaerobic respiration in yeast/ plants:
- Anaerobic respiration occurs in the absence of oxygen
- It produces less energy than its aerobic counterpart, due to the incomplete breakdown of glucose
- In yeast the overall reaction is : glucose —-> ethanol + carbon dioxide
- It involves two steps: glycolysis and fermentation
- Anaerobic respiration is characterized by the absence of the link reaction, Krebs cycle and electron transport chain
- This is because of the inability to reform NAD in these processes in the absence of oxygen
- However, glycolysis still occurs as NAD is regenerated
- Glycolysis results in the formation of both pyruvate and reduced NAD
- During the fermentation step, pyruvate (3C) undergoes decarboxylation using the enzyme pyruvate decarboxylase, resulting in the production of ethanal (2C)
- Ethanal then combines with reduced NAD to produce ethanol using the enzyme alcohol dehydrogenase
- The overall reaction is : pyruvate + NADH —-> ethanol + CO2 + NAD
- Thus, NAD is reformed and the reaction continues to take place
- 2 ATP molecules are produced
- This reaction is irreversible and leads to energy wastage
- Anaerobic respiration is useful in plants as it allows them to adapt to the continuous lack of oxygen in their environment
Anaerobic respiration in mammals:
- Also involves glycolysis and fermentation
- The difference, however, is that pyruvate directly combines with reduced NAD to produce lactate [lactic acid] using the enzyme lactate dehydrogenase
- The overall reaction is: pyruvate —-> lactate
- 2 ATP molecules are produced
- This reaction is reversible
- In fact the reverse process occurs in the liver, where lactate is converted to pyruvate to due to lactate’s toxic nature
Adaptations of rice for wet environments:
1. Cells are tolerant to high ethanol concentrations: The low oxygen concentrations present in wet environments compared to that found in the air spaces in soil means that anaerobic respiration frequently takes place, producing large volumes of ethanol
2. Stems have air spaces known as aerenchyma which facilitate the diffusion of oxygen down to the plant’s roots
3. Some plants have the capacity for elongated stem growth, exposing the leaves to oxygen on the surface
I’ll come back to try and make this presentable when I’m less sleep desprived lol. Corrections and constructive criticism are welcome as always!
0 notes
Last Seen Blogs

geminiccasa
bia

aleksey-world
Без названия

somewhereonlyino
Cultivating my own lotus pond

bloody-hydrangeas
🌺🌙💌🌙🌺

hargo-news
Hargo.co.id